Трнк и ирнк чем отличаются
ГяРНК свойственна исключительно эукариотам, её нет у прокариот («доядерные» организмы, например бактерии и вирусы). Некоторые вирусы для хранения и передачи генетической информации следующим поколениям используют РНК вместо ДНК.
ГяРНК и её производное — информационная (или матричная) РНК переносят генетическую информацию от ядерной ДНК к цитоплазме.
Количество видов гяРНК равно количеству генов, так как она служит прямой копией кодирующих последовательностей генома. В процессе транскрипции РНК с ДНК ключевую роль играет фермент РНК-полимераза II. Информационная РНК образуется в результате процессинга гяРНК, при котором происходят вырезание некодирующих участков (интронов) и склеивание кодирующих экзонов. Таким образом, в состав иРНК входят кодирующая информация соответствующих видов гяРНК, а также фланкирующий лидерный и трейлерный участки, по этой причине она значительно короче.
Транспортная РНК
Каждая молекула тРНК состоит примерно из 75 связанных между собой нуклеотидов, образующих длинную цепь. В результате взаимодействия входящих в её состав оснований тРНК имеет конформационную структуру «клеверный лист», который затем скручивается в L-форму. Очень часто в состав тРНК помимо Ц, Г, А и У входит ряд редких оснований, некоторые из которых модифицированы путём метилирования. Важная особенность тРНК: «заряженная» молекула несёт на своём 3′-конце аминокислоту, а посередине конформационной структуры «клеверный лист» находятся три специфических основания, именуемые антикодоном. Последовательность оснований в антикодоне напрямую зависит от вида аминокислоты, прикреплённой к 3′-концу.
Так, например, тРНК, антикодон которой имеет последовательность 5′-ЦЦА-3′, может нести только аминокислоту триптофан. Следует отметить, что данная зависимость лежит в основе передачи генетической информации, носителем которой выступает тРНК.
Транскрипция молекул тРНК происходит с кодирующих её последовательностей в ДНК при участии фермента РНК-полимеразы III. Различают более 40 семейств тРНК, которые, в свою очередь, подразделяют на несколько видов.
Рибосомальная РНК
Существует несколько субъединиц рРНК, которые различаются по коэффициенту седиментации (осаждения), измеряемому в единицах Сведберга (S). Данный коэффициент зависит от скорости осаждения субъединиц при центрифугировании в насыщенной водной среде.
Каждая рибосома состоит из большой и малой субъединиц. Они содержат большое количество белков, синтезированных посредством трансляции иРНК, а также РНК, которая не подвергается трансляции. Термин «рибосомальная РНК» относят именно к нетранслируемому материалу. В малой субъединице находится 18S рРНК, а в большой — 4S, 5,8S и 28S рРНК.
Траскрипция рРНК с ДНК происходит при помощи двух дополнительных РНК-полимераз. РНК-полимераза I транскрибирует 5S, 5,8S и 28S в виде одного длинного 45S-тpaнскрипта, который затем разделяется на необходимые части. Таким образом обеспечивается равное количество молекул. В организме человека в каждом гаплоидном геноме присутствует примерно 250 копий последовательности ДНК, кодирующей 45S-транскрипт. Они расположены в пяти кластерных тандемных повторах в коротких плечах хромосом 13, 14, 15, 21 и 22.
Данные участки известны как ядрышковые организаторы, так как их транскрипция и последующий процессинг 45S-транскрипта происходят внутри ядрышка.
Не менее чем в трёх кластерах хромосомы 1 существует 2000 копий 5S-pPHK гена. Их транскрипция протекает в присутствии РНК-полимеразы III снаружи ядрышка. Затем они доставляются к местам сборки рибосом при помощи рибосомальных белков.
В рРНК насчитывают около 95 псевдоуридиновых участков, образованных посредством изомеризации уридина малой ядрышковой РНК.
Малая ядрышковая РНК. Малая (низкомолекулярная) ядрышковая РНК в основном участвует в направлении или проведении модификаций оснований в рРНК и малой ядерной РНК, таких, как, например, метилирование и псевдоуридинизация. Большинство малых ядрышковых РНК находятся в интронах других генов.
Сигналраспознающая РНК. Сигналраспознающая РНК распознаёт сигнальную последовательность белков, предназначенных для экспрессии, и участвует в их переносе через цитоплазматическую мембрану.
Микро-РНК. Существует примерно 200 микро-РНК человека длиной в 22 основания, производных расщепления рибонуклеазой Н их предшественников (двухцепочечных «зашпиленных» РНК) в соответствии с инвертированными повторами. Они контролируют трансляцию структурных генов путём комплементарного связывания с З’-концами нетранслируемых участков иРНК.
Митохондриальная РНК
Митохондриальная ДНК представляет собой непрерывную петлю и кодирует 13 полипептидов, 22 тРНК и 2 рРНК (16S и 23S). Большинство генов находятся на одной (тяжёлой) цепи, однако некоторое их количество расположено и на комплементарной ей лёгкой. При этом обе цепи транскрибируются в виде непрерывных транскриптов при помощи митохондриоспецифической РНК-полимеразы. Данный фермент кодируется ядерным геном. Длинные молекулы РНК затем расщепляются на 37 отдельных видов, а мРНК, рРНК и тРНК совместно транслируют 13 мРНК. Большое количество дополнительных белков, которые поступают в митохондрию из цитоплазмы, транслируются с ядерных генов.
У пациентов с системной красной волчанкой обнаруживают антитела к снурп-белкам собственного организма. Кроме того, считают, что определённый набор генов малой ядерной РНК хромосомы 15q играет важную роль в патогенезе синдрома Прадера—Вилли (наследственное сочетание олигофрении, низкого роста, ожирения, гипотонии мышц).
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Введение в биологию. Часть №8
Оригинал взят у  caenogenesis
caenogenesis 
Тема IX
ГЕНЕТИЧЕСКАЯ ИНФОРМАЦИЯ



Тут пригодятся наши знания о белках. Мы знаем, что первичная структура белка, то есть аминокислотная последовательность, как правило, однозначно предопределяет все остальные уровни его структуры. Поэтому линейный нуклеотидный текст вполне может задавать все свойства сколь угодно сложной белковой молекулы. Тогда, однако, возникает следующий вопрос: каким образом нуклеотидный “алфавит” переводится в аминокислотный?
Пока гены считались белками, все было относительно просто.
Итак, чего можно ожидать от генетического кода? Рассуждения ученых, занявшихся этим вопросом, были следующими.
● Протеиногенных аминокислот 20, а нуклеотидов в ДНК всего 4. Значит, каждая аминокислота должна кодироваться не одним нуклеотидом, а неким их сочетанием. Примерно так, например, вводятся с помощью клавиш китайские иероглифы.
● Двоек нуклеотидов (дублетов) возможно всего 16, для кодирования всех аминокислот этого не хватит. Значит, генетический код должен быть как минимум триплетным (Gamow, Ycas, 1955).
● Троек нуклеотидов (триплетов) возможно 64, то есть их намного больше, чем аминокислот.

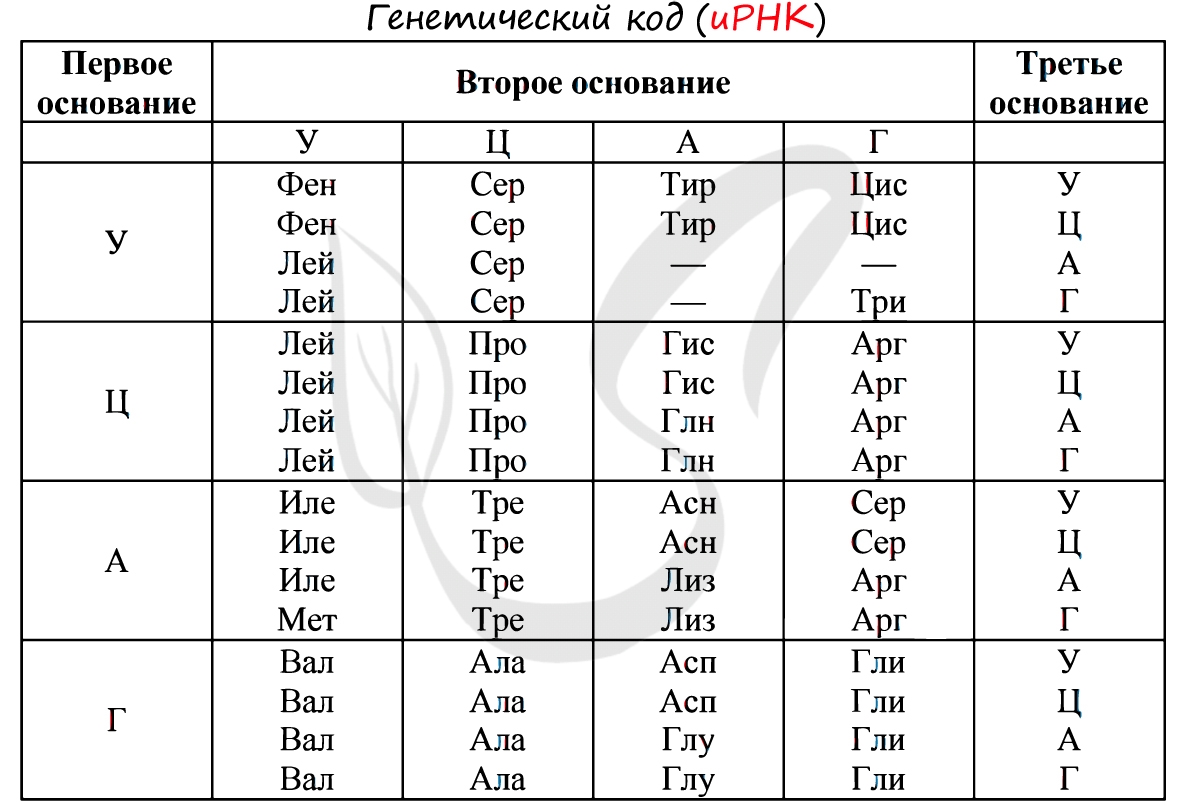
Вот так выглядит полный генетический код. Каждая тройка нуклеотидов, кодирующая определенную аминокислоту, называется кодоном. Генетический код состоит из 61 кодона, кодирующего аминокислоты, и трех стоп-кодонов, на которых синтез полипептидной цепи останавливается. Всего кодонов 64, как и предсказывал Георгий Гамов.

А с чего же тогда трансляция идет?
Молекулярно-биологические исследования быстро выявили два факта:
● Для синтеза белка совершенно необходима РНК, причем не только рибосомная, но и какая-то еще.
● У эукариот ДНК находится в ядре, в то время как синтез белка всегда идет в цитоплазме.

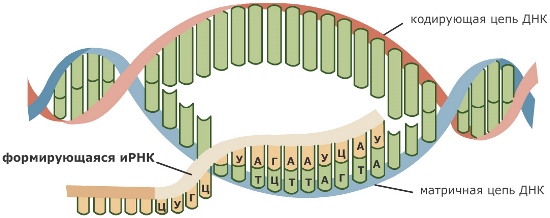
Процесс переноса информации с ДНК на РНК называется транскрипцией. Давайте посмотрим на схему транскрипции внимательно, благо всеми понятиями, которые нужны, чтобы в ней разобраться, мы теперь уже владеем. Итак, двойная спираль ДНК частично раскручивается, и фермент ДНК-зависимая РНК-полимераза ползет по одной из ее цепей от 3’-конца к 5’-концу, синтезируя комплементарную этой цепи РНК. Отметим, что синтезируемая РНК, точно так же как и вторая цепочка ДНК, антипараллельна той цепи, которой она комплементарна. Это означает, что 5’-конец и 3’-конец у нее направлены в другую сторону.


Переведем дух и поздравим себя.
Отныне мы знакомы с великой формулой ДНК→РНК→белок, которая с легкой руки Фрэнсиса Крика получила название центральной догмы молекулярной биологии. К этой формуле, конечно, есть много дополнений, но самое главное о потоке генетической информации мы теперь знаем. Информация передается с ДНК на белок через посредство РНК.
Теперь мы видим, что в таблице генетического кода не только можно, но и нужно заменить Т на У: во-первых, потому что трансляция всегда идет именно с РНК, а во-вторых, потому, что в ДНК нам придется постоянно разбираться в том, какая цепь кодирующая, а какая некодирующая (причем аминокислотной последовательности белка будет соответствовать последовательность некодирующей цепи, которая не транскрибируется). По всем этим причинам таблицу генетического кода чаще всего дают сразу в «РНКовом» варианте:

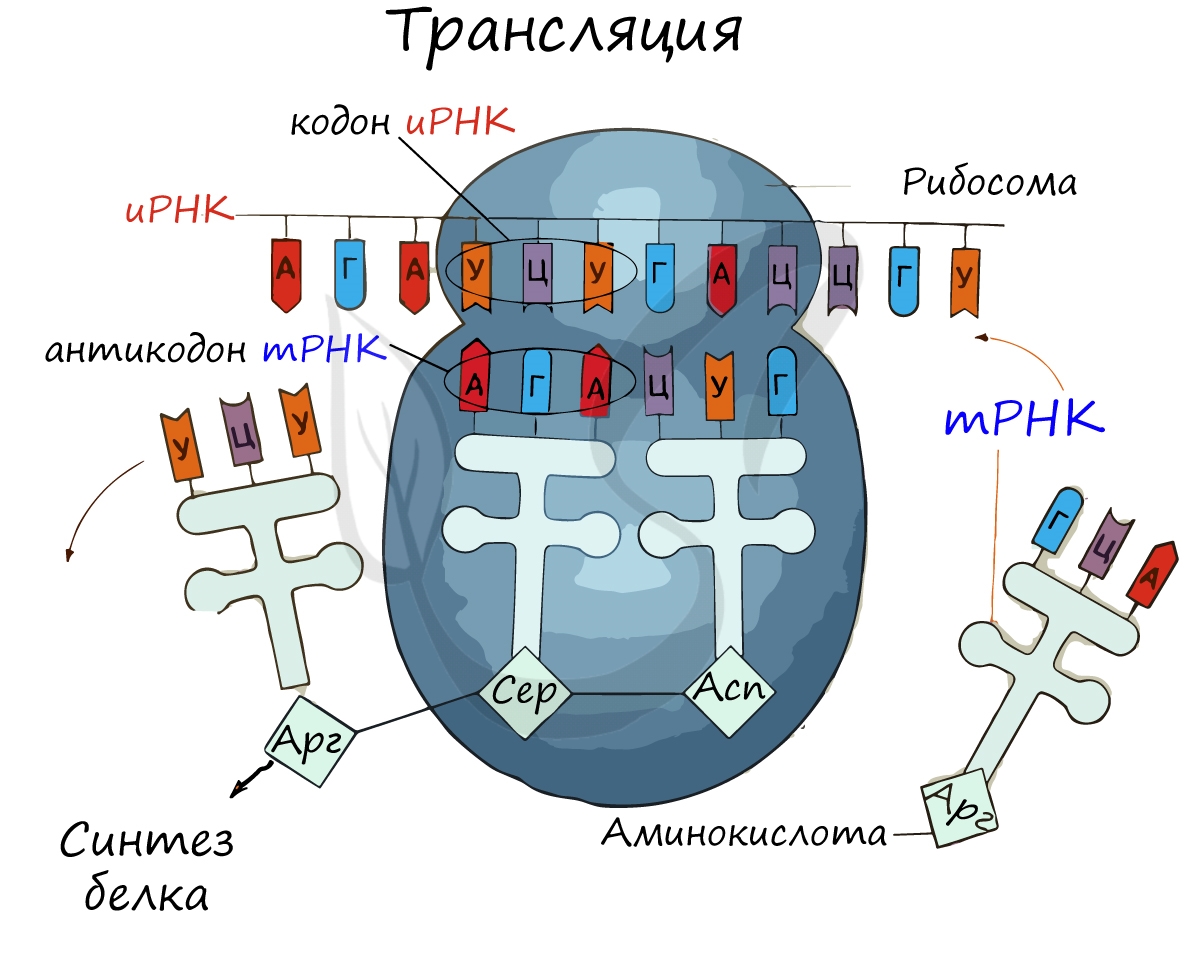
Откуда рибосома “знает”, какую аминокислоту она должна в данный момент присоединить к полипептиду? В этом ей помогает транспортная РНК (тРНК), переносящая аминокислоты. Она одноцепочечная, но имеет комплементарные спаренные участки, на которых образуются двойные спирали. Типичная конформация тРНК называется “клеверный лист”. Для каждой аминокислоты есть своя тРНК, и чаще всего не одна.




Во время трансляции любая проплывающая мимо тРНК может случайно столкнуться с тем кодоном иРНК, который в данный момент находится в активном центре рибосомы. Но свяжется она с ним только в том случае, если ее антикодон будет этому кодону комплементарен. Тогда рибосома отрежет аминокислоту от тРНК, присоединит ее к полипептидной цепочке, а сама продвинется по иРНК на шаг вперед (в сторону 3’-конца), и цикл повторится.


Теперь мы наконец можем взглянуть на самую общую схему трансляции. Здесь она очень сильно упрощена. Примерно так выглядит минимальный «сухой остаток» того, что всякому интересующемуся современной биологией стоит знать об этом процессе.
Весь процесс переноса генетической информации от ДНК через РНК к белкам называется экспрессией генов. Тут мы наконец-то сталкиваемся вплотную с понятием «ген», которое надо хоть как-то усвоить, прежде чем идти дальше.
Итак, что такое ген?
Но и способы регуляции транскрипции бывают очень разными. Неплохое представление о том, как тут все может происходить, дает вот эта относительно простая картинка:

Разберемся в ней по порядку.



Фермент, который синтезирует из мономеров новую цепь ДНК, комплементарную к имеющейся, называется ДНК-полимеразой. На самом деле в любой клетке есть несколько ДНК-полимераз, отличающихся по функциям. Но тут сразу возникает несколько проблем, которые с одним типом ферментов все равно не решить.
Во-первых, чтобы репликация стала возможна, комплементарные цепи ДНК надо как-то разделить. Для этого фермент хеликаза разрывает водородные связи между азотистыми основаниями, а фермент топоизомераза раскручивает двойную спираль ДНК, разрывая для этого ковалентные связи между нуклеотидами и тут же сшивая их заново. Последнее неизбежно, потому что двойную спираль невозможно раскрутить, не разрывая, если нам недоступны ее концы. Тут можно представить себе обыкновенный узел, концы шнурков от которого уходят куда-то в бесконечность, а нам тем не менее надо разделить шнурки, чтобы они шли параллельно и не перепутывались. Не будет другого выхода, кроме как разрезать их и потом сшить. Вот это топоизомераза и делает.
Во-вторых, ДНК-полимераза не может начать создавать новую цепь с нуля. Ей нужна затравка в виде короткой комплементарной РНК, которую синтезирует фермент праймаза. Новая ДНК может синтезироваться в виде серии фрагментов, ковалентно связанных с РНК-затравками (фрагменты Оказаки). Потом ферменты вырезают РНК, помещают на ее место комплементарные исходной цепи дезоксирибонуклеотиды, и ДНК-лигаза сшивает все это в единую цепь ДНК.


В некотором смысле биология и есть наука о поведении конвариантно редуплицирующихся структур.
Биология в лицее
Site biology teachers lyceum № 2 Voronezh city, Russian Federation
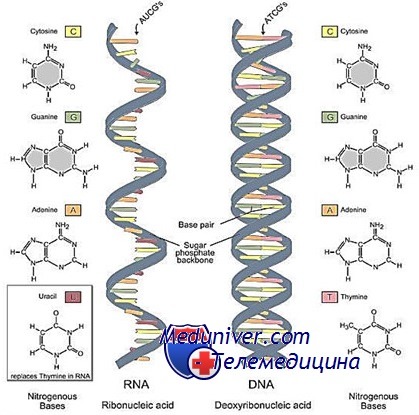
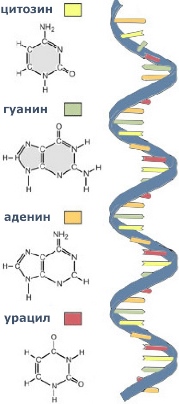 Молекула РНК (рибонуклеиновой кислоты) представляет собой неразветвленный полинуклеотид, обладающий третичной структурой. Она образована одной полинуклеотидной цепочкой, и, хотя входящие в ее состав комплементарные нуклеотиды также способны образовывать между собой водородные связи, эти связи возникают между нуклеотидами одной цепочки.
Молекула РНК (рибонуклеиновой кислоты) представляет собой неразветвленный полинуклеотид, обладающий третичной структурой. Она образована одной полинуклеотидной цепочкой, и, хотя входящие в ее состав комплементарные нуклеотиды также способны образовывать между собой водородные связи, эти связи возникают между нуклеотидами одной цепочки.
Цепи РНК значительно короче цепей ДНК. Если содержание ДНК в клетке относительно постоянно, то содержание РНК сильно колеблется. Наибольшее количество РНК в клетках наблюдается во время синтеза белка.
РНК принадлежит главная роль в передаче и реализации наследственной информации. В соответствии с функцией и структурными особенностями различают три основных класса клеточных РНК:
Мономерами в рибонуклеиновой молекуле являются нуклеотиды, состоящие из остатка фосфорной кислоты, рибозы и азотистого основания (одного из четырех: аденина, гуанина, тимина или урацила).
Информационную РНК (иРНК) называют еще иногда матричной РНК (мРНК).
Ее молекулы наиболее разнообразны по размерам, молекулярной массе (от 5х10 4 до 4х10 6 ) и стабильности. Информационные РНК составляют 2 — 5% от общего количества рибонуклеиновых кислот в клетке.
Рибосомальная РНК (рРНК) в комплексе с белками образует рибосомы — органоиды, на которых происходит сборка белковых молекул.
Рибосомальные РНК состоят из 3 — 5 тыс. нуклеотидов, имеют молекулярную массу 1 — 1,5 млн.; рРНК составляют 80 — 85% от общего содержания рибонуклеиновых кислот в клетке. Рибосомальные РНК эукариотических и прокариотических клеток отличаются по размеру и синтезируются в клетках эукариот в ядрышке, а в клетках прокариот в цитоплазме.
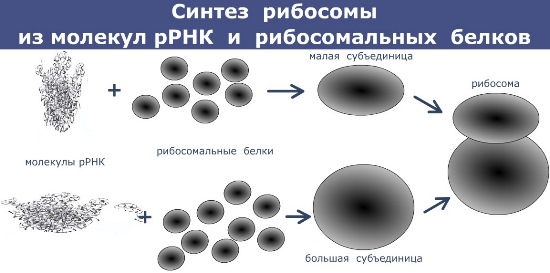
Основное значение рРНК состоит в том, что они обеспечивают первоначальное связывание иРНК и рибосомы и формируют активный центр рибосомы, в котором происходит образование пептидных связей между аминокислотами в процессе синтеза полипептидной цепи.
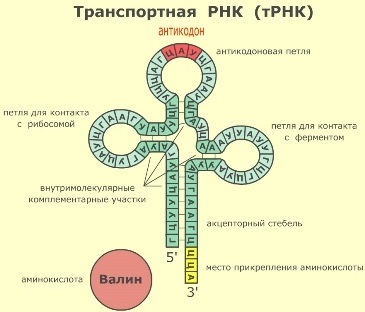 Транспортные РНК (тРНК) играют роль посредников в биосинтезе белка — они доставляют аминокислоты к месту синтеза белка, то есть на рибосомы.
Транспортные РНК (тРНК) играют роль посредников в биосинтезе белка — они доставляют аминокислоты к месту синтеза белка, то есть на рибосомы.
Молекулы тРНК содержат обычно 75 — 95 нуклеотидов; молекулярная масса 25 — 30 тыс. На долю тРНК в клетке приходится около 10% от общей массы рибонуклеиновых кислот.
У молекул тРНК имеется несколько внутримолекулярных комплементарных участков, благодаря этому их третичная структура напоминает по форме клеверный лист. У каждой тРНК есть акцепторный участок (место прикрепления транспортируемой аминокислоты), петля для контакта с рибосомой, петля для контакта с ферментом и антикодоновая петля. Антикодон — это три нуклеотида антикодоновой петли, комплементарные соответствующему кодону иРНК.
В клетке содержится около 30 видов тРНК. Каждый вид тРНК имеет характерную только для него последовательность нуклеотидов. Таким образом, конкретная тРНК может транспортировать только один вид аминокислоты, соответствующий ее антикодону.
РНК вирусов. Геном РНК-содержащих вирусов представлен двухспиральной либо односпиральной РНК. У некоторых вирусов геномные РНК сходны с иРНК эукариот и могут непосредственно транслироваться.
Сравнительная характеристика ДНК и РНК
Нуклеиновые кислоты
Строение нуклеотида
Функция в клетке
Особенность строения молекулы биополимера
1) остаток фосфорной кислоты
2) дезоксирибоза
3) азотистое основание (аденин, или гуанин, или цитозин, или тимин)
хранитель наследственной информации
1) остаток фосфорной кислоты
2) рибоза
3) азотистое основание (аденин, или гуанин, или цитозин, или урацил)
информационная, транспортная РНК принимают участие в синтезе белка
Транскрипция и трансляция

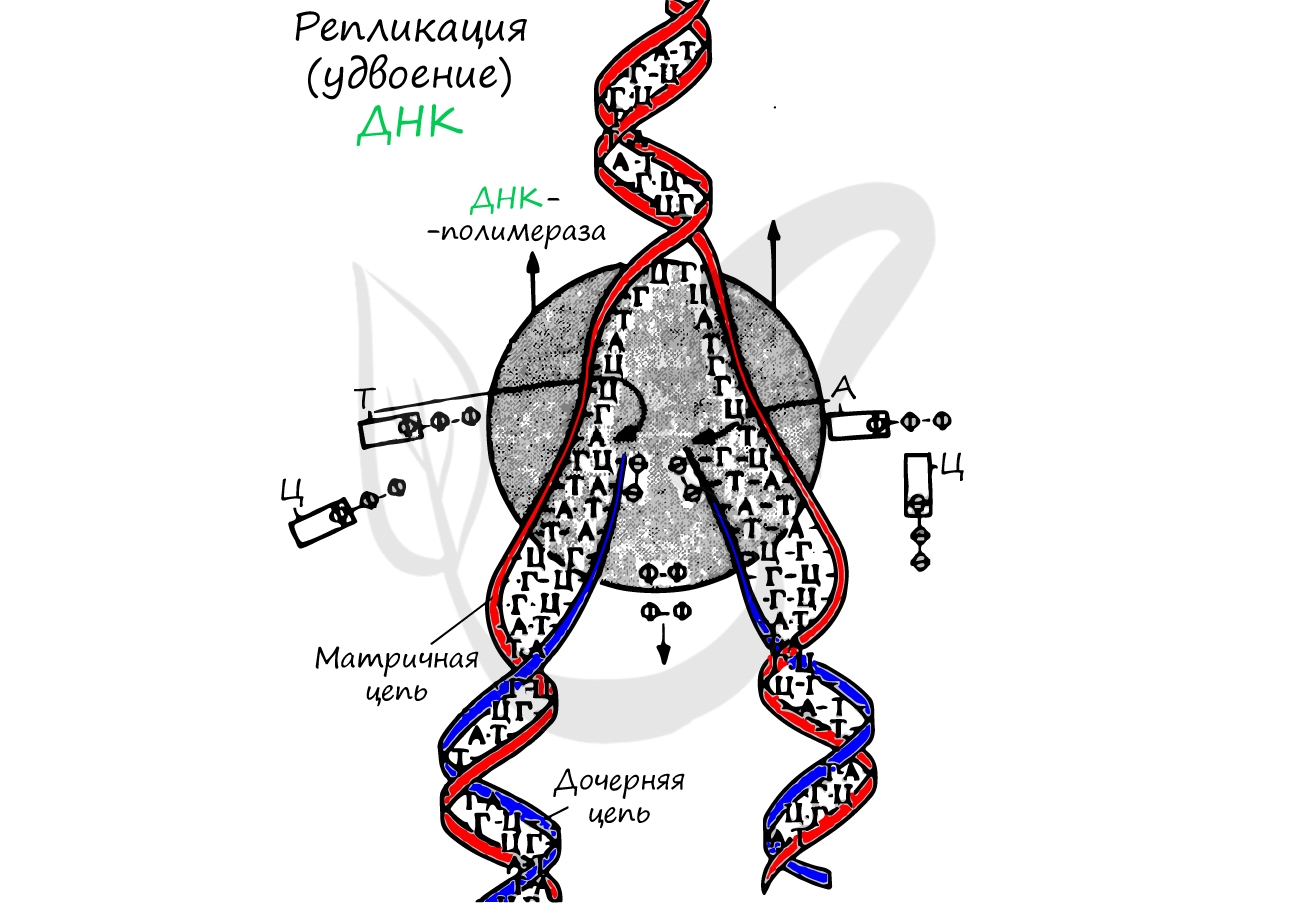
Удвоение ДНК происходит в синтетическом периоде интерфазы. При этом общее число хромосом не меняется, однако каждая из них содержит к началу деления две молекулы ДНК: это необходимо для равномерного распределения генетического материала между дочерними клетками.
Транскрпиция (лат. transcriptio — переписывание)
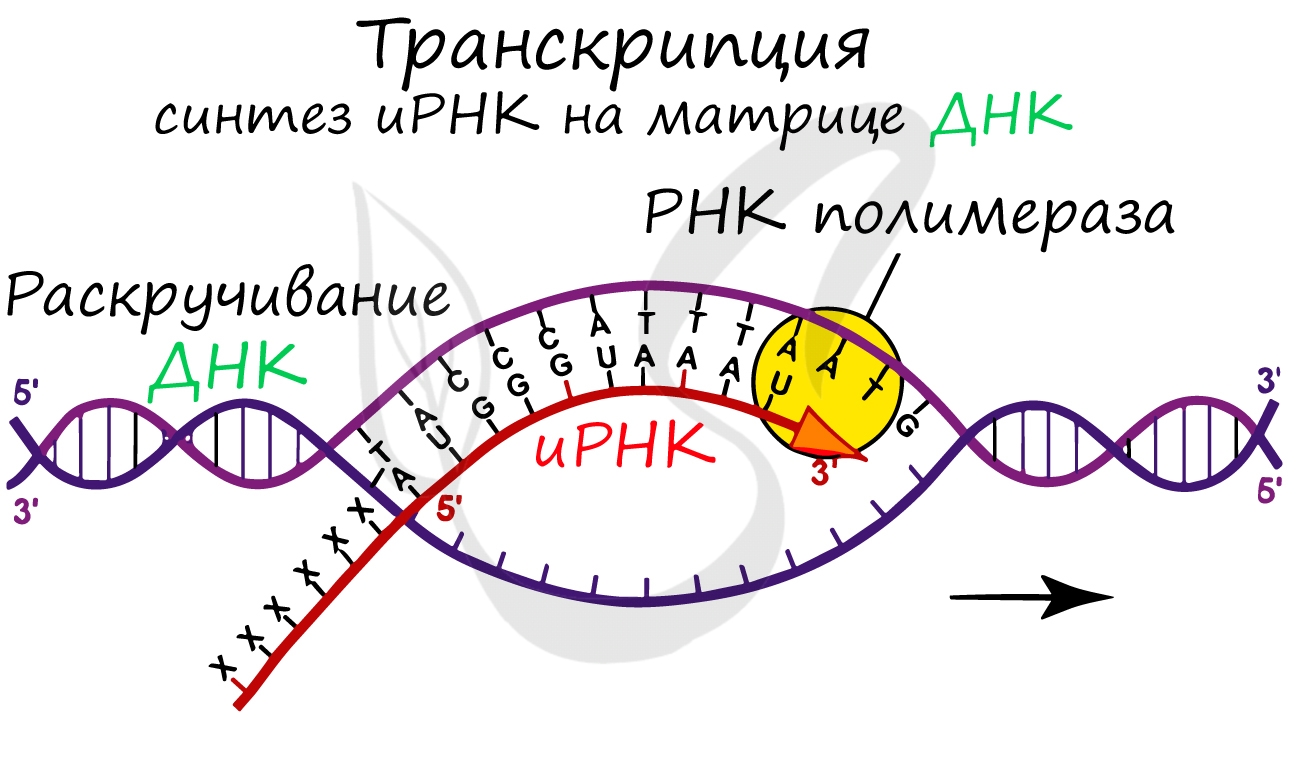
Образуется несколько начальных кодонов иРНК.
Нити ДНК последовательно расплетаются, освобождая место для передвигающейся РНК-полимеразы. Молекула иРНК быстро растет.
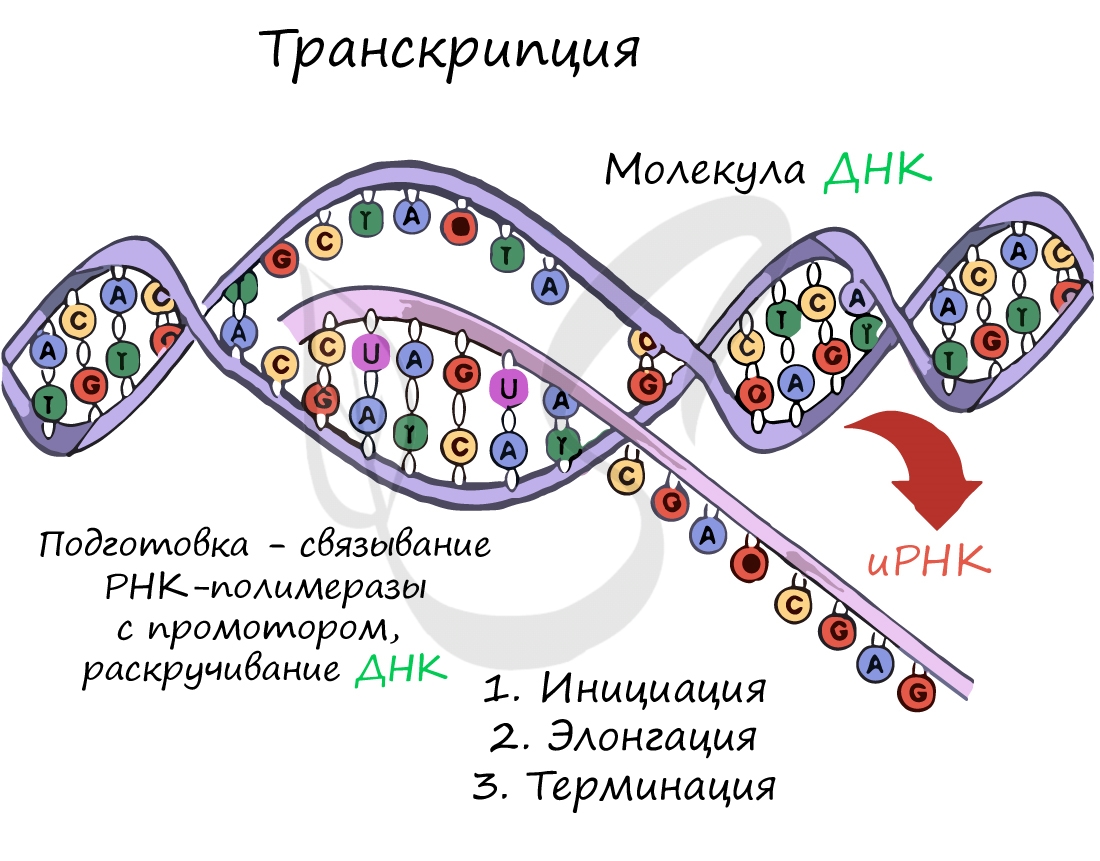
Трансляция (от лат. translatio — перенос, перемещение)
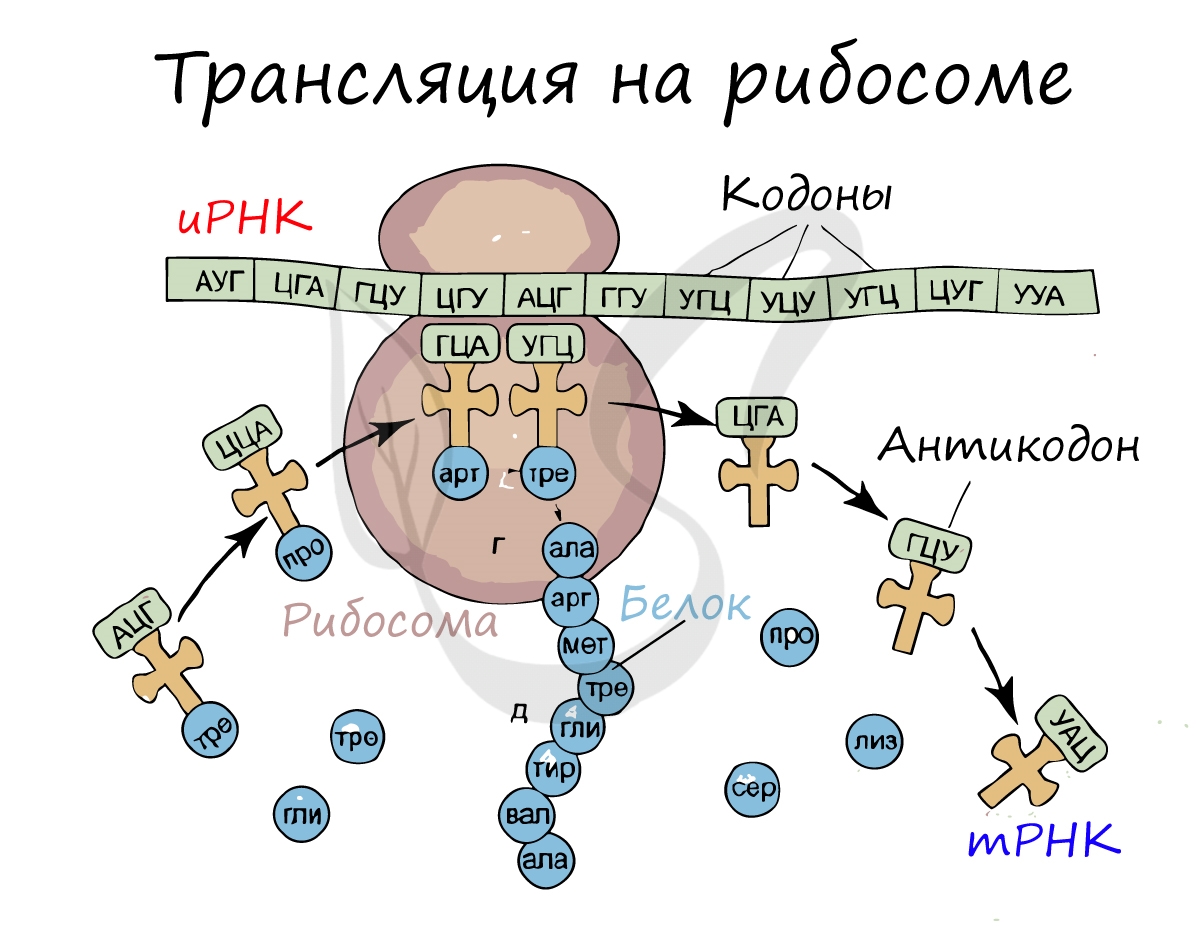
Рибосома делает шаг, и иРНК продвигается на один кодон: такое в фазу элонгации происходит десятки тысяч раз. Молекулы тРНК приносят новые аминокислоты, соответствующие кодонам иРНК. Аминокислоты соединяются друг с другом: между ними образуются пептидные связи, молекула белка растет.
Примеры решения задачи №1
Без практики теория мертва, так что скорее решим задачи! В первых двух задачах будем пользоваться таблицей генетического кода (по иРНК), приведенной вверху.
«Фрагмент цепи ДНК имеет следующую последовательность нуклеотидов: ЦГА-ТГГ-ТЦЦ-ГАЦ. Определите последовательность нуклеотидов во второй цепочке ДНК, последовательность нуклеотидов на иРНК, антикодоны соответствующих тРНК и аминокислотную последовательность соответствующего фрагмента молекулы белка, используя таблицу генетического кода»

По принципу комплементарности мы нашли вторую цепочку ДНК: ГЦТ-АЦЦ-АГГ-ЦТГ. Мы использовали следующие правила при нахождении второй нити ДНК: А-Т, Т-А, Г-Ц, Ц-Г.
Вернемся к первой цепочке, и именно от нее пойдем к иРНК: ГЦУ-АЦЦ-АГГ-ЦУГ. Мы использовали следующие правила при переводе ДНК в иРНК: А-У, Т-А, Г-Ц, Ц-Г.
Зная последовательность нуклеотидов иРНК, легко найдем тРНК: ЦГА, УГГ, УЦЦ, ГАЦ. Мы использовали следующие правила перевода иРНК в тРНК: А-У, У-А, Г-Ц, Ц-Г. Обратите внимание, что антикодоны тРНК мы разделяем запятыми, в отличие кодонов иРНК. Это связано с тем, что тРНК представляют собой отдельные молекулы (в виде клеверного листа), а не линейную структуру (как ДНК, иРНК).
Пример решения задачи №2
«Известно, что все виды РНК синтезируются на ДНК-матрице. Фрагмент цепи ДНК, на которой синтезируется участок центральной петли тРНК, имеет следующую последовательность нуклеотидов: ТАГ-ЦАА-АЦГ-ГЦТ-АЦЦ. Установите нуклеотидную последовательность участка тРНК, который синтезируется на данном фрагменте, и аминокислоту, которую будет переносить эта тРНК в процессе биосинтеза белка, если третий триплет соответствует антикодону тРНК»

Пример решения задачи №3
Длина фрагмента молекулы ДНК составляет 150 нуклеотидов. Найдите число триплетов ДНК, кодонов иРНК, антикодонов тРНК и аминокислот, соответствующих данному фрагменту. Известно, что аденин составляет 20% в данном фрагменте (двухцепочечной молекуле ДНК), найдите содержание в процентах остальных нуклеотидов.

Теперь мы украсили теорию практикой. Что может быть лучше при изучении новой темы? 🙂
© Беллевич Юрий Сергеевич 2018-2021
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Генетический код. Биосинтез белка
теория по биологии 🌿 основы генетики
Генетическая информация и генетический код
Каждый вид имеет свой собственный, отличный от других видов, набор белков. Интересно то, что белки, выполняющие идентичные функции у разных видов могут быть похожими или даже абсолютно идентичными.
У белков есть несколько состояний их структур:
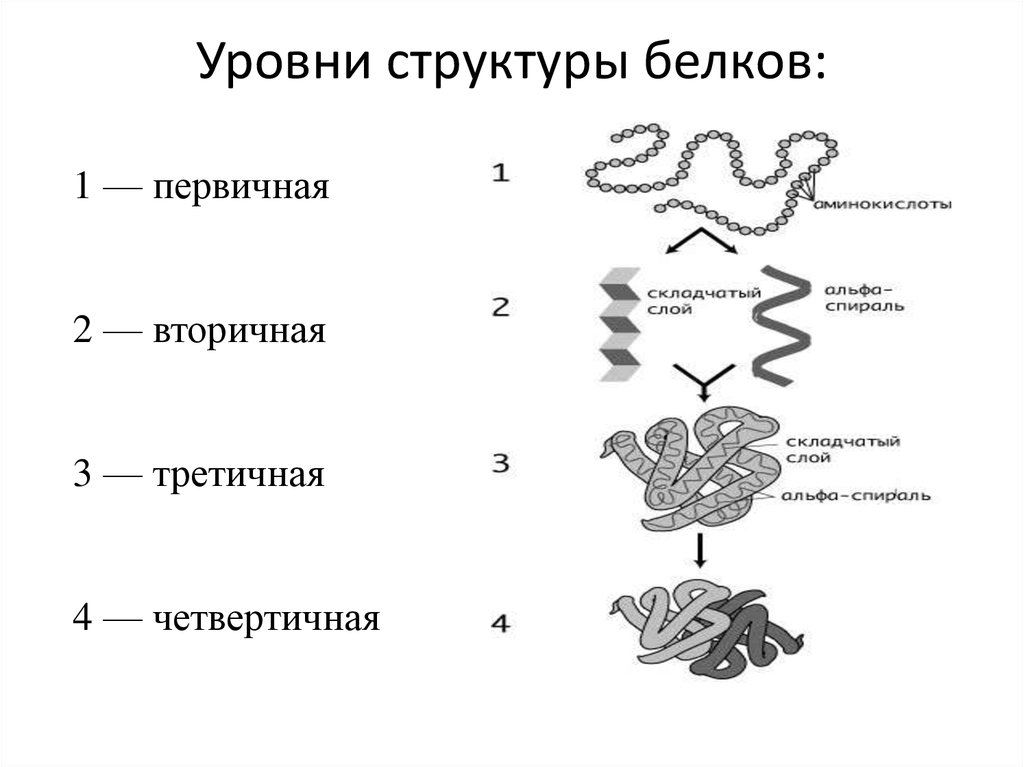
Именно первичная структура является определяющей свойства белка. Эта структура – цепь из аминокислот. Аминокислоты, в свою очередь, представляют собой ряд триплетов из нуклеотидов. Решая генетические задачи, обращаются как раз-таки к знакомой таблице:
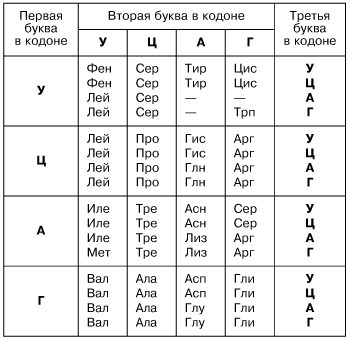
Каждая аминокислота кодируется тремя нуклеотидами, которые составляют триплет или иначе кодон. Именно последовательность нуклеотидов называется генетической информацией, а участок последовательности, в котором хранится информация о первичной структуре белка это и есть ген.
Нуклеотиды, составляющие ДНК и РНК различаются:
В состав ДНК входят:
В состав РНК входят:
Кроме того, в составе РНК (рибонуклеиновой кислоты) сахар рибоза, а ДНК (дезоксирибонуклеиновой кислоты) — дезоксирибоза. РНК — одноцепочечная, а ДНК — двухцепочечная.
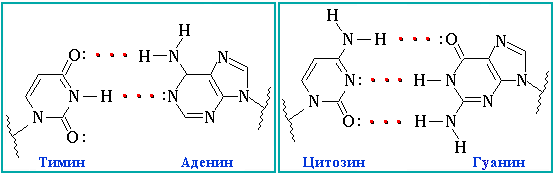
Между нуклеотидами есть водородные связи. Они могут быть как двойные, так и тройные. Нуклеотиды не могу быть связаны в случайном порядке. Для этого существует принцип комплементарности ДНК, по которому аденин одной цепи ДНК соединяется с тимином другой цепи ДНК, другая пара в ДНК – гуанин – цитозин. В РНК все аналогично, за исключением того, что вместо тимина там урацил. Между парами А-Т/А-У – две водородных связи, а между парами Ц-Г – три. На письме это обозначается чёрточками: двойная связь как знак «равно», а тройная – три горизонтальные черты.
Свойства генетического кода
Транскрипция и трансляция
Из цитологии известно, что генетическая информация у эукариотических клеток заключена в ядре в виде ДНК. Однако процесс биосинтеза белка происходит в цитоплазме на рибосомах.
Спиральная цепь ДНК при раскручивается, в это время по одной из цепочек ДНК строится комплементарная цепь. Из ядра в цитоплазму информация выходит в виде информационной РНК (иРНК). иРНК комплементарная одной из цепей ДНК. Этот процесс переписывания называется транскрипцией. Полученная цепь практически идентичная другой цепи ДНК, за исключением того, что вместо тимина там урацил.В процессе участвует специальный фермент РНК-полимераза.
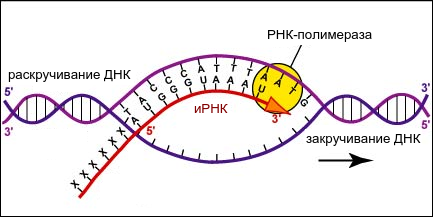
Теперь в ядре есть цепочка, которая уже начала процесс биосинтеза. Как говорилось выше, процесс ассимиляции идет на рибосомах. иРНК выходит в цитоплазму через поры ядерной мембраны
тРНК по форме напоминает лист клевера, а по принципу работы – штамп. На него, прямо как чернила, наслаиваются кодоны.
В цитоплазме начинается процесс трансляции, то есть перевод последовательности нуклеотидов информационной РНК в последовательность аминокислот белка.
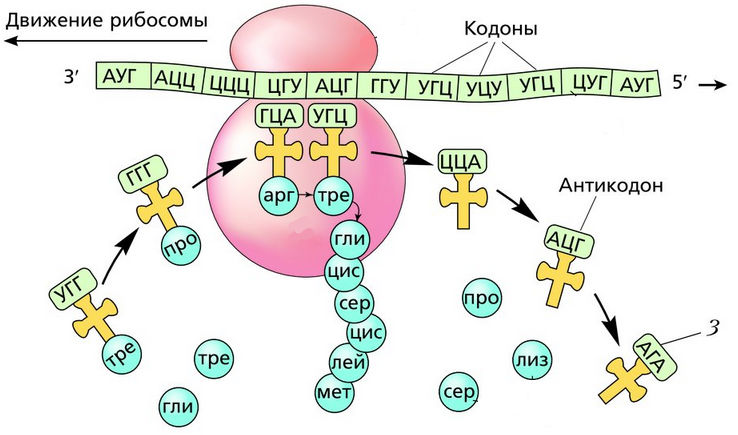
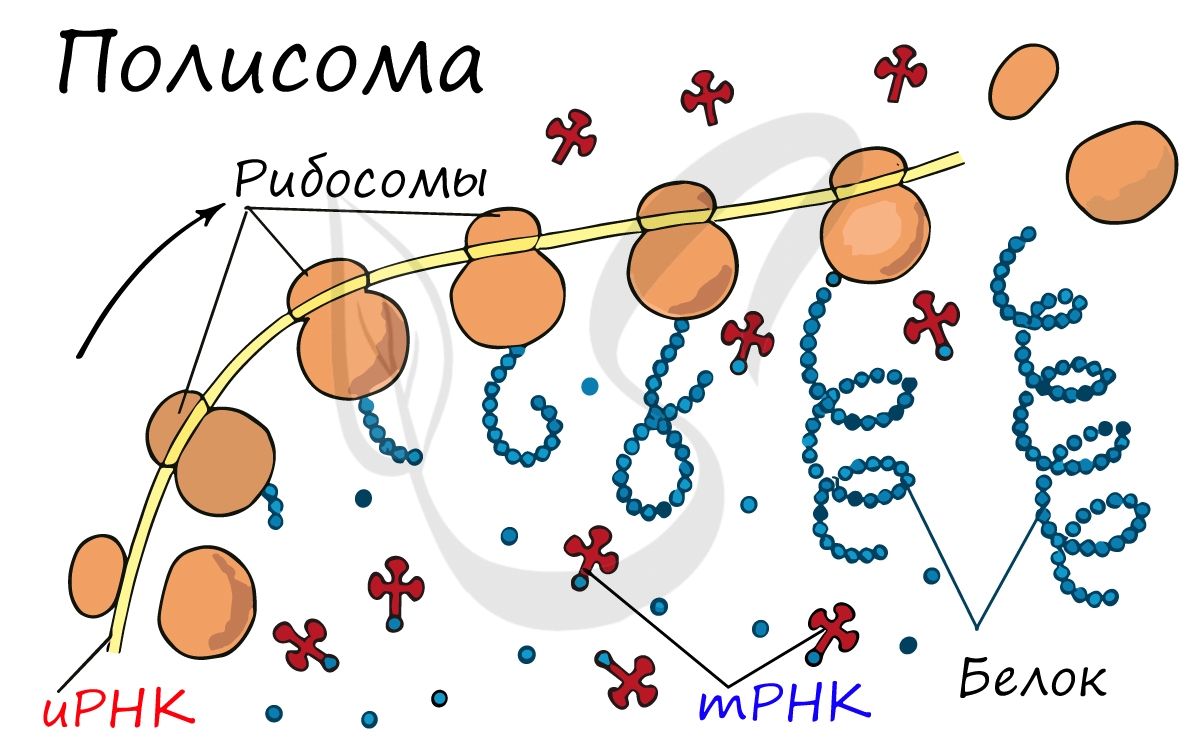
Рибосома захватывает стартовый конец цепи иРНК. Затем она начинает двигаться по цепи, одна остановка рибосомы происходит на 6-ти нуклеотидах. В это время молекула тРНК, на которых есть триплет аминокислоты «подлетает» к цепи, в месте, где находится рибосома. За время остановки рибосомы транспортная РНК успевает распознать свою пару на цепи иРНК, которая называется антикодоном. Тогда тРНК «ставит свой штамп», оставляя на цепи свой кодон. Между нуклеотидами образуются водородные связи. Так нарастает новая цепь. На одной информационной РНК работает сразу много рибосом, поэтому работа идет очень быстро. Совокупность рибосом, синтезирующих на одной иРНК, называется полисомой.
По окончанию процесса биосинтеза, цепочка отсоединяется от рибосомы и принимает свою природную структуру: вторичную, третичную или четвертичную.
pазбирался: Надежда | обсудить разбор | оценить
В современной генетической инженерии часто применняют технологии, связанные с гомологичной рекомбинацией ДНК непосредственно в живом объекте. Один из примеров – система CRE-Lox P. Lox P – это последовательность нуклеотидов в ДНК фага Р1. Она состоит из 34 нуклеотидов. В середине располагается несимметричная последовательность из 8 нуклеотидов (показана серой стрелкой на рисунке). По краям располагаются так называемые палиндромные последовательности из 13 нуклеотидов (выделены на рисунке как пунктирные блоки). Они симметричны (чтобы в этом убедиться, достаточно прочитать обе последовательность от 5´- конца к 3´- концу). Именно эти палиндромные участки узнаёт особый фермент, вызывающий рекомбинацию, который обозначают CRE. Будем в дальнейшем называть этот фермент рекомбиназой CRE. Для того, чтобы состоялась рекомбинация, два сайта Lox P должны расположиться параллельно друг другу. Рекомбиназа CRE узнает эти сайты, внесет в ДНК разрезы в определённых местах, а затем 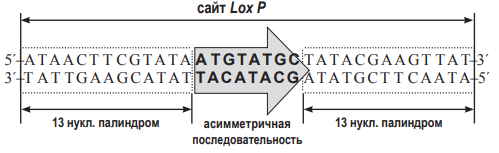 соединит по-новому две нити ДНК (т.е. произойдет рекомбинация). Аналогично работает и другая система гомологичной рекомбинации – Flp-FRT, обнаруженная у пекарских дрожжей. Сайт FRT – это последовательность ДНК, которую узнает свой фермент гомологичной рекомбинации – флиппаза (Flp).
соединит по-новому две нити ДНК (т.е. произойдет рекомбинация). Аналогично работает и другая система гомологичной рекомбинации – Flp-FRT, обнаруженная у пекарских дрожжей. Сайт FRT – это последовательность ДНК, которую узнает свой фермент гомологичной рекомбинации – флиппаза (Flp). 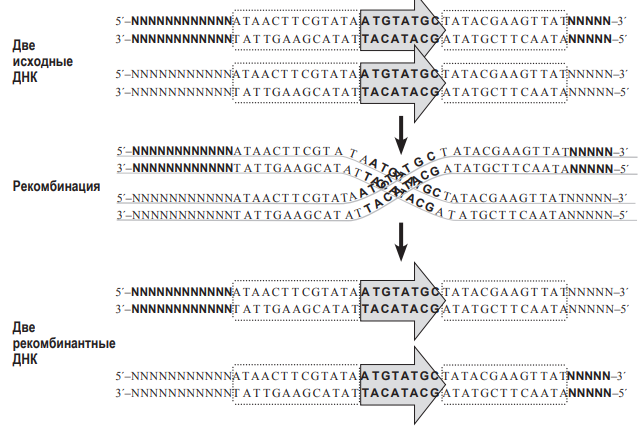 При рекомбинации две молекулы ДНК должны ориентироваться параллельно друг другу сайтами FRT, и только в этом случае произойдёт рекомбинация. Заметим, что флиппаза Flp узнает только свою последовательность FRT, но не может работать с сайтами Lox P, а рекомбиназа CRE узнает только свои сайты Lox P, но не работает с сайтами FRT. Предварительное доказательство (лемма) к задаче 9 (5 баллов). 1. Докажем, что при гомологичной рекомбинаци по «перевёрнутым» (инвертированным) повторам происходит «переворот» последовательности ДНК, находящейся между повторами. Для этого нарисуем молекулу ДНК и условно обозначим на ней буквами несколько точек.
При рекомбинации две молекулы ДНК должны ориентироваться параллельно друг другу сайтами FRT, и только в этом случае произойдёт рекомбинация. Заметим, что флиппаза Flp узнает только свою последовательность FRT, но не может работать с сайтами Lox P, а рекомбиназа CRE узнает только свои сайты Lox P, но не работает с сайтами FRT. Предварительное доказательство (лемма) к задаче 9 (5 баллов). 1. Докажем, что при гомологичной рекомбинаци по «перевёрнутым» (инвертированным) повторам происходит «переворот» последовательности ДНК, находящейся между повторами. Для этого нарисуем молекулу ДНК и условно обозначим на ней буквами несколько точек.  Затем «изогнём» молекулу так, чтобы повторы, обозначенные стрелками, встали параллельно друг другу. После обмена участками и «распрамления» окажется, что центральная часть между повторами «перевернулась».
Затем «изогнём» молекулу так, чтобы повторы, обозначенные стрелками, встали параллельно друг другу. После обмена участками и «распрамления» окажется, что центральная часть между повторами «перевернулась». 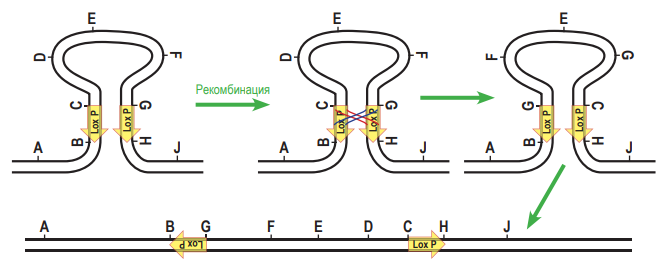 2. Докажем, что при гомологичной рекомбинаци по прямым повторам происходит образование кольцевой ДНК, при этом из линейной последовательности ДНК «удаляется» участок, находящейся между повторами. Для этого используем тот же приём: нарисуем молекулу ДНК и условно обозначим на ней буквами несколько точек. Только в этом случае для того, чтобы прямые повторы встали параллельно друг другу, придётся хитроумно изогнуть молекулу так, чтобы от конца одного из повторов (точка С) шли точки D, E, F, а потом начинался новый повтор (в точке G). После рекомбинации точки С и G поменяются местами, и в результате получится кольцевая ДНК (C, D, E, F, G) и линейный участок (A, B, H, J). Будем считать, что кольцевая ДНК как бы «исчезает» (не может реплицироваться в клетке).
2. Докажем, что при гомологичной рекомбинаци по прямым повторам происходит образование кольцевой ДНК, при этом из линейной последовательности ДНК «удаляется» участок, находящейся между повторами. Для этого используем тот же приём: нарисуем молекулу ДНК и условно обозначим на ней буквами несколько точек. Только в этом случае для того, чтобы прямые повторы встали параллельно друг другу, придётся хитроумно изогнуть молекулу так, чтобы от конца одного из повторов (точка С) шли точки D, E, F, а потом начинался новый повтор (в точке G). После рекомбинации точки С и G поменяются местами, и в результате получится кольцевая ДНК (C, D, E, F, G) и линейный участок (A, B, H, J). Будем считать, что кольцевая ДНК как бы «исчезает» (не может реплицироваться в клетке). 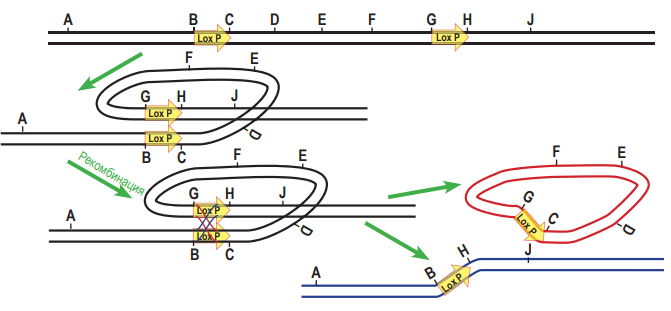 А. Поскольку после 35S-промотора на той же цепи ДНК располагается кодирующая часть гена DsRed, клетки должна светиться красным светом. Б. Рекомбиаза CRE узнаёт последовательнсоти LoxP. Если повторы расположены инвертированно, то произойдёт «переворот» последовательности ДНК, расположенной между повторами. Таким образом, после рекомбинации конструкция будет выглядеть следущим образом:
А. Поскольку после 35S-промотора на той же цепи ДНК располагается кодирующая часть гена DsRed, клетки должна светиться красным светом. Б. Рекомбиаза CRE узнаёт последовательнсоти LoxP. Если повторы расположены инвертированно, то произойдёт «переворот» последовательности ДНК, расположенной между повторами. Таким образом, после рекомбинации конструкция будет выглядеть следущим образом: 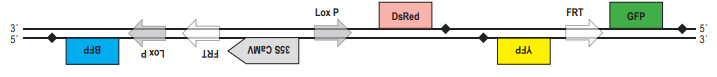 Свечение клеток изменится, поскольку после промотора на той же цепи ДНК окажется гена BFP, обестпечивающий синее свечение клеток. В. При рекомбинации по прямым повторам происходит потеря участка ДНК, расположенного между ними. Из двух повторов остаётся только один. Таким образом, после рекомбинации по сайтам FRT конструкция будет выглядеть следующим образом:
Свечение клеток изменится, поскольку после промотора на той же цепи ДНК окажется гена BFP, обестпечивающий синее свечение клеток. В. При рекомбинации по прямым повторам происходит потеря участка ДНК, расположенного между ними. Из двух повторов остаётся только один. Таким образом, после рекомбинации по сайтам FRT конструкция будет выглядеть следующим образом: 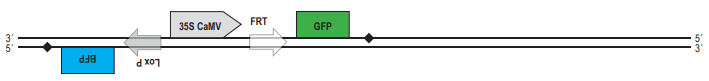 Клетки будут светиться зелёным светом за счёт того, что под промотором оказалась кодирующая последовательность гена GFP. Г. После действия рекомбиназы CRE те последовательности, на которые может действовать флип паза Flp, «перевернулись», и вместо прямых стали инвертрованными. После рекомбинации участок между ними также должен «перевернуться»:
Клетки будут светиться зелёным светом за счёт того, что под промотором оказалась кодирующая последовательность гена GFP. Г. После действия рекомбиназы CRE те последовательности, на которые может действовать флип паза Flp, «перевернулись», и вместо прямых стали инвертрованными. После рекомбинации участок между ними также должен «перевернуться»: 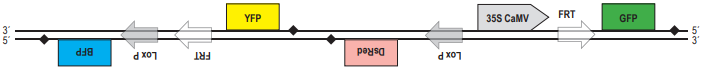 В этом случае клетки также будут светиться зелёным светом за счёт того, что под промотором оказалась кодирующая последовательность гена GFP.
В этом случае клетки также будут светиться зелёным светом за счёт того, что под промотором оказалась кодирующая последовательность гена GFP.
pазбирался: Надежда | обсудить разбор | оценить
pазбирался: Надежда | обсудить разбор | оценить
Сначала найдём место расщепления плазмиды рестриктазой BglII:  Таких участков оказывается два. В результате расщепления из плазмиды выщепляется короткий фрагмент:
Таких участков оказывается два. В результате расщепления из плазмиды выщепляется короткий фрагмент:  Остаётся укороченная линейная ДНК, содержащая интактный ген устойчивости к ампицилину и расщеплённый ген устойчивости к эритромицину.
Остаётся укороченная линейная ДНК, содержащая интактный ген устойчивости к ампицилину и расщеплённый ген устойчивости к эритромицину.  При сшивании липких концов ДНК-лигазой наиболее часто будут соединяться концы этой молекулы и образовываться кольцо длиной 4163 нуклеотида. Такая ДНК будет сообщать клеткам устойчивость к ампицилину и не даст устойчивости к эритромицину. Второй фрагмент из-за небольшой длины не может замкнуться в кольцо. Второй вариант лигирования приводит к сшиванию липких концов двух фрагментов. Он происходит примерно в 10 раз реже, а после сшивки вторая пара липких концов скорее всего также, как и исходный фрагмент замкнётся в кольцо. Таких колец из пары фрагментов может образоваться 4 вида: димеры большого фрагмента в двух разных ориентациях (правый конец с левым концом второго фрагмента и левый конец с правым концом второго фрагмента или правый с правым и левый с левым) и соединения большого и малого фрагмента в двух разных ориентациях (вариант исходной плазмиды и инверсия малого фрагмента). Из них только в варианте исходной плазмиды восстанавливается устойчивость к эритромицину. Линейная молекула, образованная сшиванием двух фрагментов, может присоединить ещё один фрагмент с ещё в 10 раз меньшей частотой. Такие фрагменты в дальнейшем будут циклизоваться в плазмиды трёх размеров: из трёх больших фрагментов, из двух больших и одного малого и одного большого и двух малых. Три малых фрагмента дадут короткую последовательность, которая не сможет замкнуться в кольцо и существовать в клетке. В каждом размерном классе будет несколько вариантов с разной ориентацией фрагментов. Только в одном из них восстановится ген устойчивости к эритромицину: правый конец большого фрагмента соединяется с левым концом малого фрагмента, а правый конец малого фрагмента – с левым концом второго большого фрагмента, а оставшиеся концы двух больших фрагментов соединяются с образованием кольцевой плазмиды длиной 8363 пары нуклеотидов. Доля таких молекул будет менее 1% всех плазмид. Вероятность образования плазмид из 4 и более фрагментов ещё на порядок ниже и их обнаружение при данном числе полученных трансформированных клеток нереально. А. Так как расщепление рестриктазой не затрагивает ген устойчивости к ампицилину, все клетки, в результате трансформации получившие любую плазмиду, будут устойчивы к ампицилину и вырастут на среде с этим антибиотиком. Таким образом из 33506 выросших колоний плазмиду получили 578, выросших на ампицилине. Эффективность трансформации представляет долю трансформированных клеток от общего их числа, т.е. 573 : 51366 × 100% = 1.12% Б. На эритромицине могут вырасти только те клетки, в которые попали плазмиды, в которых в результате лигирования восстановится последовательность нуклеотидов в гене устойчивости к этому антибиотику, расщеплённому рестриктазой. Остальные плазмиды, полученные по приведённой методике, будут содержать либо ген с выщепленным коротким фрагментом, что приведёт либо к утрате стартового кодона (если обозначенный зелёным цветом кодон является стартовым), либо к сдвигу рамки считывания (т.к. число удалённых нуклеотидов не кратно трём), либо, при инверсии короткого фрагмента, к появлению стоп-кодонов т.е. прекращению синтеза белка. Таким образом большинство полученных плазмид не обеспечат устойчивости к эритромицину. В. Рост на эритромицине могут обеспечить только плазмиды, несущие восстановленную последовательность гена устойчивости. Такие плазмиды могли образоваться из одного большого и одного малого фрагмента (4200 пар, исходная плазмида)) или из двух больших и одного малого (8363 пары, начало и конец гена из разных копий большого фрагмента). Г. Получается 1 размер из одного большого фрагмента, два размерных класса из двух фрагментов и три размерных класса из трёх фрагментов, то есть 6 размерных классов. (В реальности различить по длине плазмиды, отличающиеся на длину малого фрагмента, т.е. менее чем на 0,5%, невозможно. Поэтому в эксперименте, например на электрофореграмме, будут видны лишь три размерных класса, соответствующие 1, 2 или 3 копиям большого фрагмента.)
При сшивании липких концов ДНК-лигазой наиболее часто будут соединяться концы этой молекулы и образовываться кольцо длиной 4163 нуклеотида. Такая ДНК будет сообщать клеткам устойчивость к ампицилину и не даст устойчивости к эритромицину. Второй фрагмент из-за небольшой длины не может замкнуться в кольцо. Второй вариант лигирования приводит к сшиванию липких концов двух фрагментов. Он происходит примерно в 10 раз реже, а после сшивки вторая пара липких концов скорее всего также, как и исходный фрагмент замкнётся в кольцо. Таких колец из пары фрагментов может образоваться 4 вида: димеры большого фрагмента в двух разных ориентациях (правый конец с левым концом второго фрагмента и левый конец с правым концом второго фрагмента или правый с правым и левый с левым) и соединения большого и малого фрагмента в двух разных ориентациях (вариант исходной плазмиды и инверсия малого фрагмента). Из них только в варианте исходной плазмиды восстанавливается устойчивость к эритромицину. Линейная молекула, образованная сшиванием двух фрагментов, может присоединить ещё один фрагмент с ещё в 10 раз меньшей частотой. Такие фрагменты в дальнейшем будут циклизоваться в плазмиды трёх размеров: из трёх больших фрагментов, из двух больших и одного малого и одного большого и двух малых. Три малых фрагмента дадут короткую последовательность, которая не сможет замкнуться в кольцо и существовать в клетке. В каждом размерном классе будет несколько вариантов с разной ориентацией фрагментов. Только в одном из них восстановится ген устойчивости к эритромицину: правый конец большого фрагмента соединяется с левым концом малого фрагмента, а правый конец малого фрагмента – с левым концом второго большого фрагмента, а оставшиеся концы двух больших фрагментов соединяются с образованием кольцевой плазмиды длиной 8363 пары нуклеотидов. Доля таких молекул будет менее 1% всех плазмид. Вероятность образования плазмид из 4 и более фрагментов ещё на порядок ниже и их обнаружение при данном числе полученных трансформированных клеток нереально. А. Так как расщепление рестриктазой не затрагивает ген устойчивости к ампицилину, все клетки, в результате трансформации получившие любую плазмиду, будут устойчивы к ампицилину и вырастут на среде с этим антибиотиком. Таким образом из 33506 выросших колоний плазмиду получили 578, выросших на ампицилине. Эффективность трансформации представляет долю трансформированных клеток от общего их числа, т.е. 573 : 51366 × 100% = 1.12% Б. На эритромицине могут вырасти только те клетки, в которые попали плазмиды, в которых в результате лигирования восстановится последовательность нуклеотидов в гене устойчивости к этому антибиотику, расщеплённому рестриктазой. Остальные плазмиды, полученные по приведённой методике, будут содержать либо ген с выщепленным коротким фрагментом, что приведёт либо к утрате стартового кодона (если обозначенный зелёным цветом кодон является стартовым), либо к сдвигу рамки считывания (т.к. число удалённых нуклеотидов не кратно трём), либо, при инверсии короткого фрагмента, к появлению стоп-кодонов т.е. прекращению синтеза белка. Таким образом большинство полученных плазмид не обеспечат устойчивости к эритромицину. В. Рост на эритромицине могут обеспечить только плазмиды, несущие восстановленную последовательность гена устойчивости. Такие плазмиды могли образоваться из одного большого и одного малого фрагмента (4200 пар, исходная плазмида)) или из двух больших и одного малого (8363 пары, начало и конец гена из разных копий большого фрагмента). Г. Получается 1 размер из одного большого фрагмента, два размерных класса из двух фрагментов и три размерных класса из трёх фрагментов, то есть 6 размерных классов. (В реальности различить по длине плазмиды, отличающиеся на длину малого фрагмента, т.е. менее чем на 0,5%, невозможно. Поэтому в эксперименте, например на электрофореграмме, будут видны лишь три размерных класса, соответствующие 1, 2 или 3 копиям большого фрагмента.)
pазбирался: Надежда | обсудить разбор | оценить
По принципу комплементарности строим