Немного эндокринной гинекологии

Давно хочу начать этот цикл статей по эндокринной гинекологии.
Правильнее было бы начать с параметров «нормального» менструального цикла. Эту информацию про себя должна знать каждая женщина.
Менструальный цикл (МЦ) – это цикл созревания и овуляции фолликула в яичнике с целью обеспечения зачатия, а при отсутствии беременности с исходом в менструальное кровотечение. МЦ очень ярко отражает гормональное, соматическое и психологическое здоровье женщины!
Менструальное кровотечение характеризуется такими параметрами как: длительность, наличие/отсутствие боли, обильность.
Длительность менструального кровотечения 2-7 дней. Причём чётко надо выделять: кровотечение и кровомазанье, кровомазанье не относится к менструальному кровотечению – это отдельный симптом и его, конечно, надо учитывать.
Кровомазанье – это такой тип кровянистого отделяемого, на который за 1 сутки (не учитывая условий гигиены) требуется не более 1 гигиенической прокладки, и длится оно не более 2 дней. Кровомазанье может быть в норме, но такое, чтобы в сумме с менструальным кровотечением они не составили более 8 дней!
Боль при менструации не должна длиться более 1 суток и нарушать привычный образ жизни.
Менархе – это начало первого менструального кровотечения в жизни девочки, оно не должно быть позже 16 лет, и не раньше 10,5 лет.
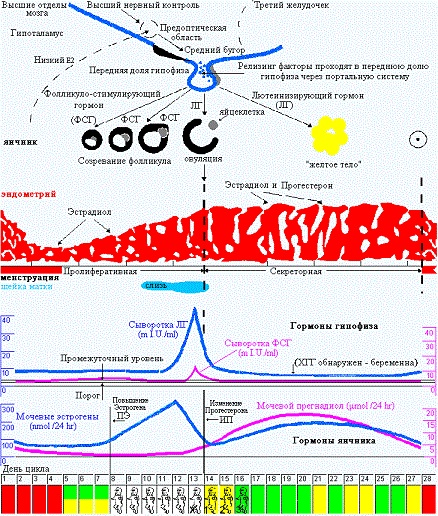
Теперь поговорим о тех гормональных изменениях, которые вызывают такую цикличность в организме. В начале, в истоке всего, стоит «цикличность работы репродуктивной системы», она «заложена» в головном мозге – гипоталамусе, из которого выходят пульсирующие импульсы стимулирующие выработку главных половых гормонов гипофиза: ФСГ (фолликулостимулирующего гормона), стимулирующего в яичнике синтез эстрогенов в 1 фазу МЦ и овуляцию и ЛГ (лютеинизирующего гормона), гормона, готовящего организм к беременности и сохраняющего беременность и стимулирующего выработку прогестерона во 2 фазу цикла. Если беременность не наступила – выработка в яичниках эстрогенов/прогестерона снижается и это является сигналом для гипоталамуса/ гипофиза к очередному циклу выработки ФСГ/ЛГ, и так «по кругу».
Это я очень примитивно описала эту связь головного мозга и яичников, конечно, она гораздо объёмнее и сложнее). На рисунке 1 (а) хорошо показаны колебания гормонов в одном цикле, выраженные в истинных единицах измерения. Нормы половых гормонов здесь не привожу, об этом отдельно, при описании нарушений. А на рисунке 1 (б) представлена «циклическая закономерность» репродуктивной системы.
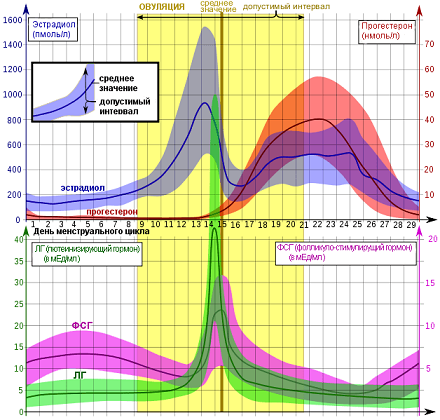
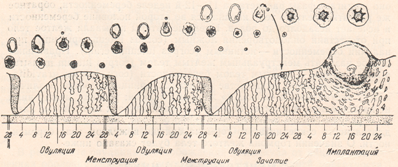
На рисунке 2 очень наглядно представлен рост фолликула, график базальной температуры и изменения эндометрия – внутреннего слоя матки в разные фазы цикла, красным – указана менструация, особенно мне нравится в этой схеме, что «детками» (8-16 день цикла) указаны дни возможного зачатия, указан «высший нервный контроль» МЦ и предоптическая область гипоталамуса, где расположено аркуатное ядро из которого исходят основные циклические импульсы к началу МЦ, указано на каком этапе ФСГ, а на каком ЛГ (главные «головные» половые гормоны) распределяют «роли» в регуляции роста фолликула, показаны пики гормонов как стимул к овуляции, указано влияние эстрогенов и прогестерона на эндометрий.
Теперь об ощущениях, которые могут сопровождать женщину в течение МЦ.
Женщины с пластичной психикой действительно чувствую перепады уровня половых гормонов в течение цикла на эмоциональном уровне. Я уже не говорю о физиологических проявлениях.
Во время менструации, как правило, (если цикл будет овуляторный) идёт повышение настроения и «прилив сил», если цикл будет неовуляторным, то к последнему дню кровотечения и на следующий день может отмечаться снижение настроения, так как при отсутствии доминантного фолликула несколько «падает» уровень эстрогенов в крови.
Далее, по окончанию менструации, настроение становится ровным примерно до 20 дня цикла (при 28 дневном МЦ), яркий пик «прилива сексуальности, сексуального желания, эмоционального подъёма» может быть в «основные» 3-4 дня овуляции, это 13-16 день цикла. 2 фаза цикла, после овуляции, в «здоровом цикле» эмоциональными всплесками не сопровождается.

В следующих статьях на эту тему я напишу о разных видах нарушений менструального цикла с позиции эндокринологии, о контрацепции и не только.
Регуляция менструального цикла
Функционирование женской репродуктивной системы — это уникальный и многоэтапный процесс, с помощью которого представляется возможным наступление беременности, вынашивание и рождение ребенка. К тому же изменения, происходящие циклически, обеспечивают нормальный уровень репродуктивных гормонов, а, соответственно, и постоянство внутренней среды организма.

Стаж работы более 22 года.

Что такое нормальный менструальный цикл?
Менструальный цикл — это постоянные изменения в женской репродуктивной системе, имеющие циклический характер, происходящие на всех уровнях ее регуляции, направленные на обеспечение наступления беременности и вынашивания ребенка. Менструальный цикл направлен на ежемесячное созревание яйцеклеток, а также создание условий для нормальной имплантации оплодотворенной яйцеклетки в эндометрий с последующим прогрессированием беременности.
Уровни регуляции менструального цикла
Регуляция цикла женщины и уровни менструального цикла – это многогранный процесс, который обеспечивает физиологическую и стабильную работу женской репродуктивной системы. Сбой хотя бы одно элемента уровня регуляции приводит каскадом в дисбаланс работу остальных звеньев этой цепи.
Уровни и схема регуляции менструального цикла.
Регуляция менструального цикла — взаимозависимая система. В регуляции менструального цикла участвуют пять уровней:

Фазы менструального цикла: гормоны
На уровне яичников можно выделить две фазы:
На уровне эндометрия можно выделить несколько гормонально зависимых фаз:
Если беременность не наступила, то гормоны в конце цикла падают, фаза секреции переходит в очередную фазу десквамации и цикл повторяется снова. Заново продуцируются гормоны по циклу.

Способы регуляции менструального цикла
Гормональная регуляция овариально-менструального цикла является сложным и многоэтапным процессом. Однако, достижения современной медицины и гинекологии в частности дают возможность регулировать этот сложный каскад реакций. Это стало возможным путем применения экзогенно гормональных средств.

Гормоны, принимающие участие в регуляции менструального цикла
Гормональная регуляция менструационного цикла осуществляется путем взаимозависимого синтеза гормонов на всех уровнях регуляции цикла.
Гормоны менструального цикла:
Гормональная регуляция менструального цикла – это созависимая система, в которой некорректная работа одного звена нарушает работу всей системы.
Данная статья не может быть использована для постановки диагноза, назначения лечения и не заменяет прием врача.
Менструальный цикл. Научный взгляд
АДГ – антидиуретический гормон
АКТГ – кортиколиберин
аРГ-Гн – агонист рилизинг-гормона гонадотропинов
ЛГ – лютеинизирующий гормон
ОП – оксипрогестерон
РГ-Гн – рилизинг-гормон гонадотропинов
СТГ – соматолиберин
СЭФР – сосудистый эндотелиальный фактор роста
ТТГ – тиреотропный гормно (тиролиберин)
ФСГ – фолликулостимулирующий гормон
ФФР – фибропластический фактор роста
Нормальный менструальный цикл
Менструации – это кровянистые выделения из половых путей женщины, периодически возникающие в результате отторжения функционального слоя эндометрия в конце двухфазного менструального цикла.
Комплекс циклических процессов, которые происходят в женском организме и внешне проявляются менструациями, называется менструальным циклом. Менструация начинается как реакция на изменение уровня стероидов, продуцируемых яичниками.
Клинические признаки нормального менструального цикла
Длительность менструального цикла в активный репродуктивный период женщины составляет в среднем 28 дней. Продолжительность цикла от 21 до 35 дней считается нормальным. Большие промежутки наблюдаются во время полового созревания и климакса, что может быть проявлением ановуляции, которая может отмечаться в это время наиболее часто.
Обычно менструация длится от 3 до 7 дней, количество теряемой крови незначительно. Укорочение или удлинение менструального кровотечения, а также появление скудных или обильных менструаций может служить проявлением ряда гинекологических заболеваний.
Характеристики нормального менструального цикла:
Длительность: 28±7 дней;
Длительность менструального кровотечения: 4±2 дня;
Объем кровопотери при менструации: 20-60 мл*;
Средняя потеря железа: 16 мг
* 95 процентов здоровых женщин с каждой менструацией теряют менее 60 мл крови. Кровопотеря более 60-80 мл сочетается со снижением уровня гемоглобина, гематокрита и сывороточного железа.
Физиология менструального кровотечения:
Непосредственно перед менструацией развивается выраженный спазм спиральных артериол. После дилатации спиральных артериол начинается менструальное кровотечение. Поначалу адгезия тромбоцитов в сосудах эндометрия подавляется, но затем, по мере транссудации крови, поврежденные концы сосудов запечатываются внутрисосудистыми тромбами, состоящими из тромбоцитов и фибрина. Через 20 ч после начала менструации, когда большая часть эндометрия уже отторглась, развивается выраженный спазм спиральных артериол, за счет чего и достигается гемостаз. Регенерация эндометрия начинается через 36 ч после возникновения менструации, несмотря на то, что отторжение эндометрия еще полностью не закончено.
Регуляция менструального цикла является сложным нейрогуморальным механизмом, который осуществляется с участием 5 основных звеньев регуляции. К ним относятся: кора головного мозга, подкорковые центры (гипоталамус), гипофиз, половые железы, периферические органы и ткани (матка, маточные трубы, влагалище, молочные железы, волосяные фолликулы, кости, жировая ткань). Последние носят название органов-мишеней, благодаря наличию рецепторов, чувствительных к действию гормонов, которые вырабатывает яичник на протяжении менструального цикла. Цитозолрецепторы – рецепторы цитоплазмы, обладают строгой специфичностью к эстрадиолу, прогестерону, тестостерону, в то время как ядерные рецепторы могут быть акцепторами таких молекул, как инсулин, глюкагон, аминопептиды.
Рецепторы к половым гормонам обнаружены во всех структурах репродуктивной системы, а также в центральной нервной системе, коже, жировой и костной ткани и молочной железе. Свободная молекула стероидного гормона захватывается специфическим цитозолрецептором белковой природы, образующийся комплекс транслоцируется в ядро клетки. В ядре возникает новый комплекс с ядерным белковым рецептором; этот комплекс связывается с хроматином, регулирующим транскрипцию мРНК, участвующим в синтезе специфического тканевого белка. Внутриклеточный медиатор – циклическая аденозинмонофосфорная кислота (цАМФ) регулирует метаболизм в клетках ткани-мишени в соответствии с потребностями организма в ответ на воздействие гормонов. Основная масса стероидных гормонов (около 80% находится в крови и транспортируется в связанном виде. Транспорт их осуществляется специальными белками – стероидсвязывающими глобулинами и неспецифическими транспортными системами (альбуминами и эритроцитами). В связанном виде стероиды неактивны, поэтому глобулины, альбумины и эритроциты можно рассматривать как своеобразную буферную систему, контролирующую доступ стероидов к рецепторам клеток-мишеней.
Циклические функциональные изменения, происходящие в организме женщины, условно можно разделить на изменения в системе гипоталамус-гипофиз-яичники (яичниковый цикл) и матке, в первую очередь в ее слизистой оболочке (маточный цикл).

Наряду с этим, как правило, происходят циклические сдвиги во всех органах и системах женщины, в частности, в ЦНС, сердечно-сосудистой системе, системе терморегуляции, обменных процессах и т.д.
Гипоталамус
Гипоталамус – часть головного мозга, расположенная над зрительным перекрестом и образующая дно III желудочка. Это старый и стабильный компонент центральной нервной системы, общая организация которого мало менялась в процессе эволюции человека. Структурно и функционально гипоталамус связан с гипофизом. Выделяют три гипоталамические области: переднюю, заднюю и промежуточную. Каждая область образована ядрами — скоплениями тел нейронов определенного типа.
Помимо гипофиза гипоталамус влияет на лимбическую систему (миндалевидное тело, гиппокамп), таламус, мост. Указанные отделы также прямо или опосредованно влияют на гипоталамус.
Гипоталамус секретирует либерины и статины. Этот процесс регулируют гормоны, замыкающие три петли обратной связи: длинную, короткую и ультракороткую. Длинную петлю обратной связи обеспечивают циркулирующие половые гормоны, связывающиеся с соответствующими рецепторами в гипоталамусе, короткую: гормоны аденогипофиза, ультракороткую: либерины и статины. Либерины и статины регулируют активность аденогипофиза. Гонадолиберин стимулирует секрецию ЛГ и ФСГ, кортиколиберин – АКТГ, соматолиберин (СТГ), тиролиберин (ТТГ). Помимо либеринов и статинов в гипоталамусе синтезируются антидиуретический гормон и окситоцин. Эти гормоны транспортируются в нейрогипофиз, откуда попадают в кровь.

В отличие от капилляров других областей мозга капилляры воронки гипоталамуса фенестрированные. Именно они образуют первичную капиллярную сеть воротной системы.
В 70-80-х гг. была выполнена серия экспериментальных работ на обезьянах, которые позволили выявить различия функции нейросекреторных структур гипоталамуса приматов и грызунов. У приматов и человека аркуатные ядра медиобазального гипоталамуса являются единственным местом образования и выделения РГ-ЛГ, ответственного за гонадотропную функцию гипофиза. Секреция РГ-ЛГ генетически запрограммирована и происходит в определенном пульсирующем ритме с частотой примерно один раз в час. Этот ритм получил название цирхорального (часового). Область аркуатных ядер гипоталамуса получила название аркуатного осциллятора. Цирхоральный характер секреции РГ-ЛГ был подтвержден путем прямого определения его в крови портальной системы ножки гипофиза и яремной вены у обезьян и в крови у женщин с овулятор-ным циклом.
Гормоны гипоталамуса
Рилизинг-гормон ЛГ выделен, синтезирован и подробно описан. Выделить и синтезировать фоллиберин до настоящего времени не удалось. РГ-ЛГ и его синтетические аналоги обладают способностью стимулировать выделение ЛГ и ФСГ передней долей гипофиза, поэтому в настоящее время принят один термин для гипоталамических гонадотропных либеринов – рилизинг-гормон гонадотропинов (РГ-Гн).
Гонадолиберин стимулирует секрецию ФСГ и ЛГ. Это декапептид, секретируемый нейронами ядра воронки. Гонадолиберин секретируется не постоянно, а в импульсном режиме. Он очень быстро разрушается протеазами (период полуразрушения составляет 2—4 мин), поэтому его импульсация должна быть регулярной. Частота и амплитуда выбросов гонадолиберина меняются на протяжении менструального цикла. Для фолликулярной фазы характерны частые колебания небольшой амплитуды уровня гонадолиберина в сыворотке крови. К концу фолликулярной фазы частота и амплитуда колебаний возрастают, а затем снижаются на протяжении лютеиновой фазы.
Гипофиз
В гипофизе выделяют две доли: переднюю – аденогипофиз и заднюю – нейрогипофиз. Нейрогипофиз имеет нейрогенное происхождение и представляет продолжение воронки гипоталамуса. Нейрогипофиз кровоснабжается из нижних гипофизарных артерий. Аденогипофиз развивается из эктодермы кармана Ратке, поэтому состоит из железистого эпителия и не имеет прямой связи с гипоталамусом. Синтезируемые в гипоталамусе либерины и статины попадают в аденогипофиз через особую воротную систему. Это основной источник кровоснабжения аденогипофиза. Кровь в воротную систему преимущественно поступает через верхние гипофизарные артерии. В области воронки гипоталамуса они образуют первичную капиллярную сеть воротной системы, из нее формируются воротные вены, которые входят в аденогипофиз и дают начало вторичной капиллярной сети. Возможен обратный ток крови через воротную систему. Особенности кровоснабжения и отсутствие гематоэнцефалического барьера в области воронки гипоталамуса обеспечивают двустороннюю связь между гипоталамусом и гипофизом. В зависимости от окрашивания гематоксилином и эозином секреторные клетки аденогипофиза делят на хромофильные (ацидофильные) и базофильные (хромофобные). Ацидофильные клетки секретируют СТГ и пролактин, базофильные – ФСГ, ЛГ, ТТГ, АКТГ
Гормоны гипофиза
В аденогипофизе образуются СТГ, пролактин, ФСГ, ЛГ, ТТГ и АКТГ. ФСГ и ЛГ регулируют секрецию половых гормонов, ТТГ — секрецию тиреоидных гормонов, АКТГ — секрецию гормонов коры надпочечников. СТГ стимулирует рост, обладает анаболическим действием. Пролактин стимулирует рост молочных желез во время беременности и лактацию после родов.
ЛГ и ФСГ синтезируются гонадотропными клетками аденогипофиза и играют важную роль в развитии яичниковых фолликулов. По структуре относятся к гликопротеинам. ФСГ стимулирует рост фолликула, пролиферацию гранулезных клеток, индуцирует образование рецепторов ЛГ на поверхности клеток гранулезы. Под влиянием ФСГ увеличивается содержание ароматаз в зреющем фолликуле. ЛГ стимулирует образование андрогенов (предшественников эстрогенов) в тека-клетках, совместно с ФСГ способствует овуляции и стимулирует синтез прогестерона в лютеинизированных клетках гранулезы овулировавшего фолликула.
Секреция ЛГ и ФСГ непостоянна и модулируется яичниковыми гормонами, особенно эстрогенами и прогестероном.
Таким образом, низкий уровень эстрогенов оказывает подавляющий эффект на ЛГ, в то время как высокий стимулирует его производство гипофизом. В поздней фолликулярной фазе уровень сывороточных эстрогенов достаточно высок, положительный эффект обратной связи утраивается, что способствует образованию преовуляторного пика ЛГ. И, наоборот, при терапии комбинированными контрацептивами уровень эстрогенов в сыворотке крови находится в пределах, определяющих отрицательную обратную связь, что приводит к снижению содержания гонадотропинов.
Механизм положительной обратной связи приводит к повышению в рецепторах концентрации и продукции РГ-Гн.
В противоположность эффекту эстрогенов, низкий уровень прогестерона имеет положительную реакцию обратной связи на секрецию ЛГ и ФСГ гипофизом. Такие состояния существуют непосредственно перед овуляцией и приводят к выбросу ФСГ. Высокий уровень прогестерона, который отмечается в лютеиновой фазе, уменьшает гипофизарную продукцию гонадотропинов. Малое количество прогестерона стимулирует высвобождение гонадотропинов на уровне гипофиза. Отрицательный эффект обратной связи прогестерона проявляется путем уменьшения производства РГ-Гн и снижения чувствительности к РГ-Гн на уровне гипофиза. Положительный эффект обратной связи прогестерона происходит на гипофиз и включает в себя повышенную чувствительность к РГ-Гн. Эстрогены и прогестерон не являются единственными гормонами, воздействующими на секрецию гонадотропинов гипофизом. Таким же эффектом обладают гормоны ингибин и активин. Ингибин подавляет гипофизарную ФСГ секрецию, активин ее стимулирует.
Пролактин — это полипептид, состоящий из 198 аминокислотных остатков, синтезируемый лактотропными клетками аденогипофиза. Секреция пролактина контролируется дофамином. Он синтезируется в гипоталамусе и тормозит секрецию пролактина. Пролактин оказывает многообразное действие на организм женщины. Его основная биологическая роль — рост молочных желез и регуляция лактации. Он обладает также жиромобилизующим эффектом и оказывает гипотензивное действие. Увеличение секреции пролактина является одной из частых причин бесплодия, так как повышение его уровня в крови тормозит стероидогенез в яичниках и развитие фолликулов.
Окситоцин — пептид, состоящий из 9 аминокислотных остатков. Он образуется в нейронах крупноклеточной части паравентрикулярных ядер гипоталамуса. Основными мишенями окситоцина у человека служат гладкомышечные волокна матки и миоэпителиальные клетки молочных желез.
Антидиуретический гормон (АДГ) — это пептид, состоящий из 9 аминокислотных остатков. Синтезируется в нейронах супраоптического ядра гипоталамуса. Основная функция АДГ — регуляция ОЦК, АД, осмоляльности плазмы.
Яичниковый цикл
В яичниках проходит три фазы менструального цикла:
Фолликулярная фаза:
Одним из основных моментов фолликулярной фазы менструального цикла является развитие яйцеклетки. Яичник женщины представляет собой сложный орган, состоящий из многих компонентов, в результате взаимодействия которых секретируются половые стероидные гормоны и образуется готовая к оплодотворению яйцеклетка в ответ на циклическую секрецию гонадотропинов.
Гормональная активность от преантрального до периовуляторного фолликула описана как теория «две клетки, два гонадотропина». Стероидогенез происходит в двух клетках фолликула: в тека- и гранулезных клетках. В тека-клетках ЛГ стимулирует производство андрогенов из холестерола. В гранулезных клетках ФСГ стимулирует превращение полученных андрогенов в эстрогены (ароматизация). Дополнительно к эффекту ароматизации ФСГ так же отвечает за пролиферацию гранулезных клеток. Хотя известны другие медиаторы в развитии фолликулов яичника, эта теория является основной для понимания процессов, происходящих в фолликуле яичника. Выявлено, что для нормального цикла с достаточным уровнем эстрогенов необходимы оба гормона.
Производство андрогенов в фолликулах так же может регулировать развитие преантрального фолликула. Низкий уровень андрогенов усиливает процесс ароматизации, следовательно, увеличивает производство эстрогенов, и наоборот, высокий — тормозит процесс ароматизации и вызывает атрезию фолликула. Баланс ФСГ и ЛГ необходим для раннего развития фолликула. Оптимальным условием для начальной стадии развития фолликула является низкий уровень ЛГ и высокий ФСГ, что имеет место в начале менструального цикла. Если же уровень ЛГ высокий, тека-клетки производят большое количество андрогенов, вызывая атрезию фолликулов.
Выбор доминантного фолликула
Росту фолликула сопутствует секреция половых стероидных гормонов под влиянием ЛГ и ФСГ. Эти гонадотропины защищают группу преантральных фолликулов от атрезии. Однако в норме только один из этих фолликулов развивается до преовуляторного, который затем освобождается и становится доминантным.
В отличие от уровня ФСГ, который по мере увеличения концентрации эстрадиола снижается, уровень ЛГ продолжает расти (в низких концентрациях эстрадиол тормозит секрецию ЛГ). Именно длительная эстрогенная стимуляция готовит овуляторный пик ЛГ. Одновременно с этим происходит подготовка доминантного фолликула к овуляции: под местным действием эстрогенов и ФСГ на гранулезных клетках увеличивается число рецепторов ЛГ. Выброс ЛГ приводит к овуляции, образованию желтого тела и увеличению секреции прогестерона. Овуляция происходит спустя 10—12 ч после пика ЛГ или спустя 32—35 ч после начала подъема его уровня. Обычно овулирует только один фолликул.
Во время выбора фолликула уровень ФСГ понижается в ответ на отрицательный эффект от эстрогенов, поэтому доминирующий фолликул является единственным, который продолжает свое развитие при падающем уровне ФСГ
Яичниково-гипофизарная связь является решающей при выборе доминирующего фолликула и при развитии атрезии остальных фолликулов.
Рост и развитие яйцеклетки, функционирование желтого тела происходит при взаимодействии аутокринных и паракринных механизмов. Необходимо отметить два фолликулярных гормона, играющих значительную роль в стероидогенезе, — ингибин и активин.
Ингибин представляет собой пептидный гормон, вырабатываемый гранулезными клетками растущих фолликулов, снижает продукцию ФСГ. Кроме того, влияет на синтез андрогенов в яичнике. Ингибин влияет на фолликулогенез следующим образом: уменьшая ФСГ до такого уровня, при котором развивается только доминантный фолликул.
Активин представляет собой пептидный гормон, вырабатывается в гранулезных клетках фолликулов и гипофиза. По данным некоторых авторов, активин вырабатывается также и плацентой. Активин увеличивает производство ФСГ гипофизом, усиливает процесс связывания ФСГ с гранулезными клетками.
Инсулиноподобные факторы роста
Инсулиноподобные факторы роста (ИФР-1 и ИФР-2) синтезируются в печени под влиянием гормона роста и, возможно, в гранулезных клетках фолликулов, действуют как паракрин-ные регуляторы. Перед овуляцией содержание ИФР-1 и ИФР-2 в фолликулярной жидкости повышается за счет увеличения количества самой жидкости в доминантном фолликуле. ИФР-1 участвует в процессе синтеза эстрадиола. ИФР-2 (эпидермальный) тормозит синтез стероидов в яичниках.
Овуляция:
Овуляторный пик ЛГ приводит к повышению концентрации простагландинов и активности протеаз в фолликуле. Сам процесс овуляции представляет собой разрыв базальной мембраны доминантного фолликула и кровотечение из разрушенных капилляров, окружающих тека-клетки. Изменения в стенке преовуляторного фолликула, обеспечивающие ее истончение и разрыв, происходят под влиянием фермента коллагеназы; определенную роль играют также простагландины, содержащиеся в фолликулярной жидкости, протеолитические ферменты, образующиеся в гранулезных клетках, окситопин и релаксин. В результате этого в стенке фолликула образуется небольшое отверстие, через которое медленно выходит яйцеклетка. Непосредственные измерения показали, что давление внутри фолликула во время овуляции не возрастает.
В конце фолликулярной фазы ФСГ воздействует на рецепторы ЛГ в гранулезных клетках. Эстрогены являются обязательным кофактором в этом эффекте. По мере развития доминантного фолликула продукция эстрогенов увеличивается. В итоге производство эстрогенов достаточно для достижения секреции гипофизом ЛГ, что приводит к увеличению его уровня. Повышение происходит вначале очень медленно (с 8-го по 12-й день цикла), затем быстро (после 12-го дня цикла). В течение этого времени ЛГ активирует лютеинизацию гранулезных клеток в доминантном фолликуле. Таким образом, происходит выделение прогестерона. Далее прогестерон усиливает эффект эстрогенов на секрецию гипофизом ЛГ, приводя к повышению его уровня.
Овуляция происходит в течение 36 ч после начала подъема ЛГ. Определение выброса ЛГ является одним из лучших методов, который определяет овуляцию и проводится с помощью прибора «определитель овуляции».
Периовуляторный пик ФСГ, вероятно, происходит в результате положительного эффекта прогестерона. В дополнение к повышению содержания ЛГ, ФСГ и эстрогенов, во время овуляции отмечается и повышение уровня сывороточных андрогенов. Эти андрогены выделяются в результате стимулирующего эффекта ЛГ на тека-клетки, особенно в недоминирующем фолликуле.
Увеличение содержания андрогенов оказывает воздействие на усиление либидо, подтверждая, что этот период у женщин наиболее фертильный.
Уровень ЛГ стимулирует мейоз, после того как сперматозоид попадает в яйцеклетку. При выделении ооцита из яичника в овуляцию происходит разрушение стенки фолликула. Это регулируется ЛГ, ФСГ и прогестероном, которые стимулируют активность протеолитических энзимов, таких как плазминогеновые активаторы (которые выделяют плазмин, стимулирующий активность коллагеназы) и простагландины. Простагландины не только увеличивают активность протеолитических энзимов, но так же способствуют появлению воспалительно-подобной реакции в стенке фолликула и стимулируют активность гладкой мускулатуры, что способствует выходу ооцита.
Важность простагландинов в процессе овуляции доказана исследованиями, которые определяют, что снижение выделения простагландина может привести к задержке освобождения ооцита из яичника при нормальном стероидогенезе (синдром неразвивающегося лютеинизированного фолликула— СНЛФ). Так как СНЛФ нередко является причиной бесплодия, женщинам, желающим забеременеть, рекомендуется избегать приема синтезированных ингибиторов простагландина.
Лютеиновая фаза:
Строение желтого тела
После выброса яйцеклетки из яичника в полость фолликула быстро врастают формирующиеся капилляры; гранулезные клетки подвергаются лютеинизации: увеличению в них цитоплазмы и образованию липидных включений. Гранулезные клетки и текоциты образуют желтое тело — основной регулятор лютеиновой фазы менструального цикла. Клетки, образовавшие стенку фолликула, накапливают липиды и желтый пигмент лютеин и начинают секретировать прогестерон, эстрадиол-2, ингибин. Мощная сосудистая сеть способствует поступлению гормонов желтого тела в системный кровоток. Полноценное желтое тело развивается только в тех случаях, когда в преовуляторном фолликуле образуется адекватное число гранулезных клеток с высоким содержанием рецепторов ЛГ. Увеличение размеров желтого тела после овуляции происходит в основном за счет увеличения размеров гранулезных клеток, в то время как число их не увеличивается из-за отсутствия митозов. У человека желтое тело секретирует не только прогестерон, но и эстрадиол и андрогены. Механизмы регрессии желтого тела изучены недостаточно. Известно, что лютеолитическим действием обладают простагландины.

Рис. Ультразвуковая картина «цветущего» желтого тела при беременности 6 нед. 4 дня. Режим энергетического картирования.
Гормональная регуляция лютеиновой фазы
Если беременность не наступает, происходит инволюция желтого тела. Этот процесс регулируется по механизму отрицательной обратной связи: гормоны (прогестерон и эстрадиол), секретируемые желтым телом, действуют на гонадотропные клетки гипофиза, подавляя секрецию ФСГ и ЛГ. Секрецию ФСГ подавляет также ингибин. Снижение уровня ФСГ, а также местное действие прогестерона препятствует развитию группы примордиальных фолликулов.
Существование желтого тела зависит от уровня секреции ЛГ. При его снижении, обычно через 12-16 дней после овуляции, происходит инволюция желтого тела. На его месте образуется белое тело. Механизм инволюции неизвестен. Скорее всего, она обусловлена паракринными влияниями. По мере инволюции желтого тела уровень эстрогенов и прогестерона падает, что приводит к повышению секреции гонадотропных гормонов. По мере повышения содержания ФСГ и ЛГ начинает развиваться новая группа фолликулов.
Если произошло оплодотворение, существование желтого тела и секрецию прогестерона поддерживает хорионический гонадотропин. Таким образом, имплантация эмбриона приводит к гормональным изменениям, которые сохраняют желтое тело.
Длительность лютеиновой фазы у большинства женщин постоянна и составляет примерно 14 дней.
Гормоны яичников
Сложный процесс биосинтеза стероидов завершается образованием эстрадиола, тестостерона и прогестерона. Стероидпродуцирующими тканями яичников являются клетки гранулезы, выстилающие полость фолликула, клетки внутренней теки и в значительно меньшей степени строма. Клетки гранулезы и тека-клетки синергично участвуют в синтезе эстрогенов, клетки текальной оболочки являются главным источником андрогенов, которые в незначительном количестве образуются и в строме; прогестерон синтезируется в тека-клетках и клетках гранулезы.
В яичнике в раннюю фолликулярную фазу менструального цикла секретируется 60-100 мкг эстрадиола (Е2), в лютеиновую фазу – 270 мкг, к моменту овуляции – 400-900 мкг в сутки. Около 10% Е2 ароматизируется в яичнике из тестостерона. Количество эстрона, образующегося в раннюю фолликулярную фазу, составляет 60-100 мкг, к моменту овуляции синтез его возрастает до 600 мкг в сутки. Только половина количества эстрона образуется в яичнике. Вторая половина ароматизируется в Е2. Эстриол является малоактивным метаболитом эстрадиола и эстрона.
Прогестерон образуется в яичнике в количестве 2 мг/сут в фолликулярную фазу и 25 мг/сут в лютеиновую фазу менструального цикла. В процессе метаболизма прогестерон в яичнике превращается в 20-дегидропрогестерон, обладающий сравнительно малой биологической активностью.
В яичнике синтезируются следующие андрогены: андростендион (предшественник тестостерона) в количестве 1,5 мг/сут (столько же андростендиона образуется в надпочечниках). Из андростендиона образуется около 0,15 мг тестостерона, примерно такое же количество его образуется в надпочечниках.
Краткий обзор процессов, происходящих в яичниках
Фолликулярная фаза:
ЛГ стимулирует продукцию андрогенов в тека-клетках.
ФСГ стимулирует продукцию эстрогенов в гранулезных клетках.
Наиболее развитый фолликул в середине фолликулярной фазы становится доминантным.
Увеличивающееся образование эстрогенов и ингибина в доминантном фолликуле, подавляет выделение ФСГ гипофизом.
Снижение уровня ФСГ вызывает атрезию всех фолликулов кроме доминантного.
Овуляция:
ФСГ индуцирует ЛГ рецепторы.
Протеолитические ферменты в фолликуле приводят к разрушению его стенки и освобождению ооцита.
Лютеиновая фаза:
Желтое тело образуется из гранулезных и тека-клеток, сохранившихся после овуляции.
Прогестерон, выделяемый желтым телом, является доминирующим гормоном. При отсутствии беременности лютеолизис происходит через 14 дней после овуляции.
Маточный цикл
Эндометрий состоит из двух слоев: функционального и базального. Функциональный слой меняет свою структуру под действием половых гормонов и, если беременность не наступила, отторгается во время менструации.
Пролиферативная фаза:
Началом менструального цикла считают 1-й день менструации. По окончании менструации толщина эндометрия составляет 1-2 мм. Эндометрий состоит практически только из базального слоя. Железы узкие, прямые и короткие, выстланы низким цилиндрическим эпителием, цитоплазма клеток стромы почти не различается. По мере увеличения уровня эстрадиола формируется функциональный слой: эндометрий готовится к имплантации эмбриона. Железы удлиняются и становятся извитыми. Возрастает число митозов. По мере пролиферации высота эпителиальных клеток возрастает, а сам эпителий из однорядного становится многорядным к моменту овуляции. Строма отечна и разрыхлена, в ней увеличиваются ядра клеток и объем цитоплазмы. Сосуды умеренно извиты.
Секреторная фаза:
В норме овуляция происходит на 14-й день менструального цикла. Секреторная фаза характеризуется высоким уровнем эстрогенов и прогестерона. Однако после овуляции число рецепторов эстрогенов в клетках эндометрия снижается. Пролиферация эндометрия постепенно тормозится, снижается синтез ДНК, уменьшается число митозов. Таким образом, преимущественное влияние на эндометрий в секреторную фазу оказывает прогестерон.
В железах эндометрия появляются содержащие гликоген вакуоли, которые выявляются с помощью ШИК-реакции. На 16-й день цикла эти вакуоли достаточно крупные, имеются во всех клетках и находятся под ядрами. На 17-й день ядра, оттесненные вакуолями, располагаются в центральной части клетки. На 18-й день вакуоли оказываются в апикальной части, а ядра — в базальной части клеток, гликоген начинает выделяться в просвет желез путем апокриновой секреции. Наилучшие условия для имплантации создаются на 6—7-й день после овуляции, т.е. на 20—21-й день цикла, когда секреторная активность желез максимальна.
На 21-й день цикла начинается децидуальная реакция стромы эндометрия. Спиральные артерии резко извиты, позднее вследствие уменьшения отека стромы, они видны отчетливо. Сначала появляются децидуальные клетки, которые постепенно образуют скопления. На 24-й день цикла эти скопления формируют периваскулярные эозинофильные муфты. На 25-й день образуются островки децидуальных клеток. К 26-му дню цикла децидуальная реакция становится число нейтрофилов, которые мигрируют туда из крови. Нейтрофильная инфильтрация сменяется некрозом функционального слоя эндометрия.
Менструация:
Если имплантация не произошла, железы перестают продуцировать секрет, а в функциональном слое эндометрия начинаются дегенеративные изменения. Непосредственная причина его отторжения — резкий спад содержания эстрадиола и прогестерона в результате инволюции желтого тела. В эндометрии снижается венозный отток и происходит расширение сосудов. Далее происходит сужение артерий, что приводит к ишемии и повреждению тканей и функциональной потере эндометрия. Затем происходит кровотечение из фрагментов артериол, оставшихся в базальном слое эндометрия. Менструация прекращается при сужении артерий, эндометрий восстанавливается. Таким образом, прекращение кровотечения в сосудах эндометрия отличается от гемостаза в других частях организма.
Как правило, кровотечение прекращается в результате скопления тромбоцитов и отложения фибрина, что приводит к рубцеванию. В эндометрии рубцевание может привести к потере его функциональной активности (синдром Ашермана). Для избежания этих последствий необходима альтернативная система гемостаза. Сокращение сосудов является механизмом остановки кровотечения в эндометрии. При этом рубцевание минимизировано фибринолизом, который разрушает кровяные сгустки. Позже восстановление эндометрия и образование новых кровеносных сосудов (ангиогенез) приводит к завершению кровотечения в течение 5-7 дней от начала менструального цикла.
Влияние отмены эстрогенов и прогестерона на менструацию определено четко, но остается неясной роль паракринных медиаторов. Вазоконстрикторы: простагландин F2a, эндотелии-1 и тромбоцит-активизирующий фактор (ТАФ) могут производиться в пределах эндометрия и участвовать в сокращении сосудов. Так же они способствуют началу менструации и дальнейшему контролю над ней. Эти медиаторы могут регулироваться воздействием сосудорасширяющих агентов, таких как простагландин Е2, простациклин, оксид азота, которые вырабатываются эндометрием. Простагландин F2a обладает выраженным сосудосуживающим действием, усиливает спазм артерий и ишемию эндометрия, вызывает сокращения миометрия, что, с одной стороны, уменьшает кровоток, с другой – способствует удалению отторгнутого эндометрия.
Восстановление эндометрия включает в себя железистую и стромальную регенерацию и ангиогенез. Сосудистый эндотелиальный фактор роста (СЭФР) и фибропластический фактор роста (ФФР) обнаружены в эндометрии и являются сильными ангиогенезивными агентами. Выявлено, что эстрогенпродуцированная железистая и стромальная регенерация усиливается под воздействием эпидермальных факторов роста (ЭФР). Такие факторы роста как трансформирующий фактор роста (ТФР) и интерлейкины, особенно интерлейкин-1 (ИЛ-1), имеют большое значение.
Краткий обзор процессов, происходящих в эндометрии
Менструация:
Основную роль в начале менструации играет спазм артериол.
Функциональный слой эндометрия (верхний, составляющий 75% толщины) отторгается.
Менструация прекращается вследствие спазма сосудов и восстановления эндометрия. Фибринолиз препятствует образованию спаек.
Пролиферативная фаза:
Характеризуется индуцированной эстрогенами пролиферацией желез и стромы.
Секреторная фаза:
Характеризуется индуцированной прогестероном секрецией желез.
В позднюю секреторную фазу индуцируется децидуализация.
Децидуализация представляет собой необратимый процесс. При отсутствии наступления беременности в эндометрии происходит апоптоз с последующим появлением менструации.
Итак, репродуктивная система представляет собой суперсистему, функциональное состояние которой определяется обратной афферентацией составляющих ее подсистем. Выделяют: длинную петлю обратной связи между гормонами яичника и ядрами гипоталамуса; между гормонами яичника и гипофизом; короткую петлю между передней долей гипофиза и гипоталамусом; ультракороткую между РГ-ЛГ и нейроцитами (нервными клетками) гипоталамуса.
Обратная связь у половозрелой женщины имеет как отрицательный, так и положительный характер. Примером отрицательной связи является усиление выделения ЛГ передней долей гипофиза в ответ на низкий уровень эстрадиола в раннюю фолликулярную фазу цикла. Примером положительной обратной связи является выброс ЛГ и ФСГ в ответ на овуляторный максимум содержания эстрадиола в крови. По механизму отрицательной обратной связи увеличивается образование РГ-ЛГ при снижении уровня ЛГ в клетках передней доли гипофиза.
Резюме
Гонадолиберин синтезируется нейронами ядра воронки, затем попадает в воротную систему гипофиза и поступает по ней в аденогипофиз. Секреция гонадолиберина происходит импульсивно.
Ранний этап развития группы примордиальных фолликулов не зависит от ФСГ.
По мере инволюции желтого тела снижается секреция прогестерона и ингибина и повышается уровень ФСГ.
ФСГ стимулирует рост и развитие группы примордиальных фолликулов и секрецию ими эстрогенов.
Эстрогены готовят матку к имплантации, стимулируя пролиферацию и дифференцировку функционального слоя эндометрия и вместе с ФСГ способствуют развитию фолликулов.
Согласно двухклеточной теории синтеза половых гормонов, ЛГ стимулирует синтез андрогенов в текоцитах, которые затем под влиянием ФСГ превращаются в эстрогены в гранулезных клетках.
Рост концентрации эстрадиола по механизму отрицательной обратной связи, петля
которой замыкается в гипофизе и гипоталамусе, подавляет секрецию ФСГ.
Фолликул, который будет овулировать в данном менструальном цикле, называется доминантным. В отличие от остальных фолликулов, начавших рост, он несет большее число рецепторов ФСГ и синтезирует большее количество эстрогенов. Это позволяет ему развиваться, несмотря на снижение уровня ФСГ.
Достаточная эстрогенная стимуляция обеспечивает овуляторный пик ЛГ. Он, в свою очередь, вызывает овуляцию, образование желтого тела и секрецию прогестерона.
Функционирование желтого тела зависит от уровня ЛГ. При его снижении желтое тело подвергается инволюции. Обычно это происходит на 12—16-й день после овуляции.
Если произошло оплодотворение, существование желтого тела поддерживает хорионический гонадотропин. Желтое тело продолжает секретировать прогестерон, необходимый для сохранения беременности на ранних сроках.