Фенотипы и подгруппы Ч.1 Сибирид
Так как в предыдущих постах многие жаловались, что не нашли свой фенотип, решил немного по-другому подойти к этому вопросу. Сейчас (и в последующих постах на эту тему) буду печатать описание фенотипа и его подгруппы.
Древний североазиатский тип встречается у коренных сибиряков. Сегодня насчитывает несколько тысяч представителей, но был более распространен во времена мезолита. Во время неолита северно-средиземноморские группы оттеснили его.
Сибириды характеризуются короткими, толстыми телами, короткими конечностями, низкими черепами средней длины, угловатыми, не очень плоскими лицами, выраженными скулами, умеренными монгольскими складками и скромными волосами на теле, выпуклыми носами, кожа желтовато-красноватая, светло-коричневая, волосы имеют смешанный цвет. Следы Сибиридов доходят до Западной Канады и Северной Финляндии.
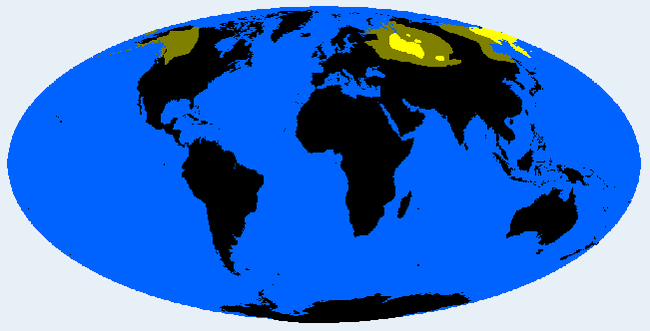


Уралид (группа Сибирид)
Западно-сибирский подтип, особенно распространенный в бассейне р. Оби среди ханты и манси. Иногда на Алтае и в Хакасии. Из-за сурового сибирского климата этот тип никогда не был очень многочисленным.
Светло-коричневая кожа, прямые или волнистые, обычно коричневые, иногда рыжие, светлые или черные волосы и смешанные глаза. Короткий, макроскликовый, эндоморфный. Мезоцефальный, шамекранический с мезорейном, умеренно выпуклый и иногда пухлый нос (особенно у женщин). Лицо у некоторых плоское. Рост волос средний, скулы и складки верхних век сильные. Губы очень тонкие.
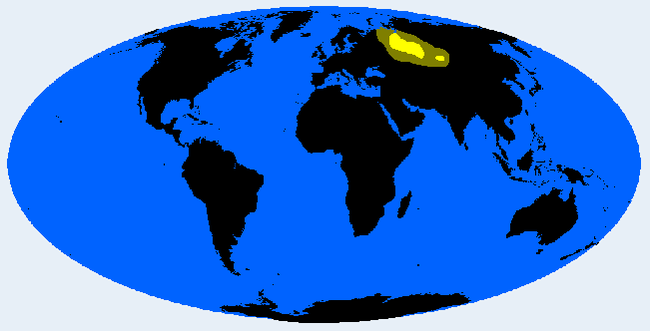


Енисей (группа Сибирид)
Сибиридный подтип бассейна реки Енисей в Сибири. Показывает некоторые американоидные особенности. Может показывать протомонголоидное состояние. Их никогда не было очень много, около 1000 особей. На более низких частотах уральцы и другие ненцы.
Желтовато-красноватая светло-коричневая кожа, прямые коричневые или черные волосы и смешанные глаза. Короче говоря, макроскопически, эктоморфно-мезоморфно. Мягко брахицефальный, шамекранический с мезоррайном, часто длинный и крючковатый нос, относительно высокий. Продолговатое лицо, косые глаза, редкая монгольская складка. Рост волос средний.
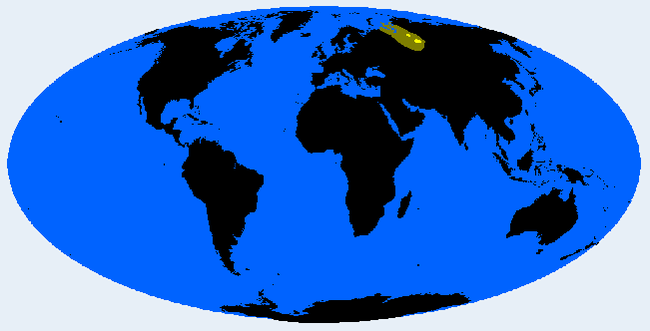


Самодийская (группа Сибирид)
Северо-западный сибиридный тип. В средние века мигрировал на полуостров Ямал и в прилегающие районы из более южного места, возможно, из Минусинского бассейна или Алтай-Саджанской области. Типично для ненцев, также в энцах, и смешано в селькупах и нганасанах. До 19 века реликтовое население существовало в горах Алтая.
Светло-желтовато-коричневая кожа с грубыми прямыми коричневыми или черными, редко светлыми волосами. Короче говоря, макроскопически, мезоморфно эндоморфно. Слегка выступающий нос. Лицо плоское и мясистое с большими выпуклыми скулами. Волосы на теле слабые, монгольская складка встречается регулярно.
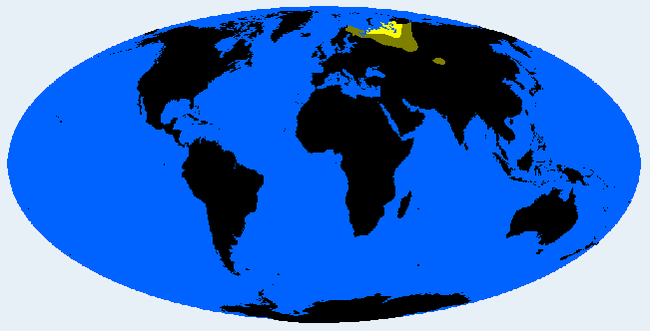


Чукчид (группа Сибирид)
Сорт дальневосточного сибирида, находящийся под влиянием эскимосов, приспособлен к одному из самых холодных обитаемых мест на Земле: арктической тундре Чукотки и Камчатки. В чистом виде у корякских и оленьих чукчей. Имеет подгруппу ительменов, которая сегодня сильно смешана с русской кровью. Иногда у прибрежных чукчей, эвенов, юкагиров, алеутов и коренных американцев северной части Тихого океана.
Светлая желтоватая / красновато-коричневая кожа с прямыми или волнистыми черными, иногда более светлыми волосами. (Скорее) короткий, эндоморфный, мускулистый, сильно макроскелированный с короткими конечностями. Мезоцефальная, иногда брахицефальная, шамо-ортокраническая. Иногда выпуклый нос (особенно у мужчин). Лицо короткое, довольно узкое и высокое, с крепкими чертами лица и массивной челюстью. Волосы на теле скудные, глаза косые, иногда с эпикантусом.
Фенотип


Феноти́п (от греческого слова phainotip — являю, обнаруживаю) — совокупность характеристик, присущих индивиду на определённой стадии развития. Фенотип формируется на основе генотипа, опосредованного рядом внешне средовых факторов. У диплоидных организмов в фенотипе проявляются доминантные гены.
Фенотип — совокупность внешних и внутренних признаков организма, приобретённых в результате онтогенеза (индивидуального развития).
Несмотря на кажущееся строгое определение, концепция фенотипа имеет некоторые неопределенности. Во-первых, большинство молекул и структур кодируемых генетическим материалом, не заметны во внешнем виде организма, хотя являются частью фенотипа. Например, именно так обстоит дело с группами крови человека. Поэтому расширенное определение фенотипа должно включать характеристики, которые могут быть обнаружены техническими, медицинскими или диагностическими процедурами. Дальнейшее, более радикальное расширение может включать приобретенное поведение или даже влияние организма на окружающую среду и другие организмы. Например, согласно Ричарду Докинзу, плотину бобров также как и их резцы можно считать фенотипом генов бобра. [1]
Фенотип можно определить как «вынос» генетической информации навстречу факторам среды. В первом приближении можно говорить о двух характеристиках фенотипа: а) число направлений выноса характеризует число факторов среды, к которым чувствителен фенотип, — мерность фенотипа; б) «дальность» выноса характеризует степень чувствительности фенотипа к данному фактору среды. В совокупности эти характеристики определяют богатство и развитость фенотипа. Чем многомернее фенотип и чем он чувствительнее, чем дальше фенотип от генотипа, тем он богаче. Если сравнить вирус, бактерию, аскариду, лягушку и человека, то богатство фенотипа в этом ряду растет.
Содержание
Историческая справка
Термин фенотип предложил датский ученый Вильгельм Иогансен в 1909 г., вместе с концепцией генотипа, чтобы различать наследственность организма от того, что получается в результате её реализации. [2] Идею о различии носителей наследственности от результата их действия можно проследить уже в работах Грегора Менделя (1865) и Августа Вейсмана. Последний различал (в многоклеточных организмах) репродуктивные клетки (гаметы) от соматических.
Факторы, определяющие фенотип
Некоторые характеристики фенотипа напрямую определяются генотипом, например цвет глаз. Другие сильно зависят от взаимодействия организма с окружающей средой — например однояйцевые близнецы могут различаться по росту, весу и другим основным физическим характеристикам, несмотря на то, что несут одни и те же гены.
Фенотипическая дисперсия
Фенотипическая дисперсия (определяемая генотипической дисперсией) является основной предпосылкой для естественого отбора и эволюции. Организм как целое оставляет (или не оставляет) потомство, поэтому естественный отбор влияет на генетическую структуру популяции опосредованно через вклады фенотипов. Без различных фенотипов нет эволюции. При этом рецессивные аллели не всегда отражаются в признаках фенотипа, но сохраняются и могут быть переданы потомству.
Фенотип и онтогенез
Факторы, от которых зависит фенотипическое разнообразие, генетическая программа (генотип), условия среды и частота случайных изменений (мутации), обобщены в следующей зависимости:
генотип + внешняя среда + случайные изменения → фенотип
Способность генотипа формировать в онтогенезе, в зависимости от условий среды, разные фенотипы называют нормой реакции. Она характеризует долю участия среды в реализации признака. Чем шире норма реакции, тем больше влияние среды и тем меньше влияние генотипа в онтогенезе. Обычно чем разнообразнее условия обитания вида, тем шире у него норма реакции.
Примеры
Иногда фенотипы в разных условиях сильно отличаются друг от друга. Так, сосны в лесу высокие и стройные, а на открытом пространстве — развесистые. Форма листьев водяного лютика зависит от того, в воде или на воздухе оказался лист. У людей все клинически определяемые признаки — рост, масса тела, цвет глаз, форма волос, группа крови и т. д. являются фенотипическими.
Фенотип лайки что такое
Войти
Авторизуясь в LiveJournal с помощью стороннего сервиса вы принимаете условия Пользовательского соглашения LiveJournal
Основные аллели окрасов собак
Наличие bb при соболином или зонарном окрасе собака будет выглядеть рыжей или коричневой, но не черной.
Для получения черных, коричневых и рыжих окрасов нужно, чтобы сочетались оба аллеля: B и Е. Их сочетание называется комплементарным. Доминирование:E>em>eb>e.
Пример:MmxMm=MM:Mm:mm=1:2:1. Мрамор х мрамор = белая: мрамор: черная. Поэтому владельцы догов предпочитают вязать мраморных собак с черными(Mm x mm = Mm:mm=1:1 мрамор х черная = мрамор: черная). У такс и колли мрамор вызывается аналогичным геном, но у них не рождается белых, не жизнеспособных щенков, но среди собак, имеющих мраморный окрас, часто встречается особи с пониженной устойчивостью к различным заболевания.
Ген Н. Он определяет доминантную окраску типа “арлекин”.
Доминантный белый окрас встречается крайне редко (южнорусская овчарка из редко лайки). От двух белых собак этого окраса возможно рождение окрашенных собак в случае гетерозиготности обоих родителей по гену W.
Маленькая буква означает, что особь несет гомозиготные рецессивные гены. ABCDEgmStrw- черная собака.
Возможные генетические формулы некоторых окрасов собак. Здесь указаны обозначения только основных генов, ответственных за данный признак.
Наследственные формулы для некоторых наиболее частых окрасок (по Н.А Ильину)
Фенотип лайки что такое
В эпоху научно-технической революции генетика является одним из наиболее актуальных, бурно развивающихся разделов биологии, всегда тесно связанным с практикой.
На основе современной генетики развивается микробиологическая промышленность, в животноводстве на генетической основе строится селекция и племенное дело, формируется генетика человека, развиваются генетические основы сохранения целостности биосферы земли и околоземного пространства.
Передача потомству признаков предыдущих поколений называется наследованием. Механизмом этой передачи служит процесс размножения, как при простом делении клеток простейших организмов и клеток тканей, так и при половом размножении, когда объединение мужских и женских половых клеток (гамет) приводит к созданию нового организма, имеющего сходство с родителями и предками,
Другие факторы внешней среды (кормление, климатические элементы и т. п.) вызывают изменения у организмов, которые не передаются потомству, т. е. не наследуются, и называются модификационной изменчивостью. Под влиянием наследственной и ненаследственной изменчивости у организмов формируется комплекс свойств, называемых фенотипической изменчивостью.
Для проведения правильного подбора родительских пар важно знать и уметь определять и выделять из фенотипической изменчивости долю влияния наследственной и ненаследственной изменчивости. Чем больше дол участия наследственности в формировании свойств и признаков организма, тем эффективнее селекционная работа.
Таким образом, механизм наследования, т. е. передачи различных признаков и свойств, действует в зависимости от молекулярного строения нуклеиновых кислот (ДНК, РНК), их генного состава. Процесс передачи этих наследственных элементов происходит размножением при делении соматических клеток и оплодотворением, при котором слияние мужских и женских гамет половых клеток приводит к образованию нового организма с удвоенным набором хромосомного аппарата. Единицей наследственности служит участок ДНК, называемый геном. Ген отца и ген матери называют аллелями гена, обусловливающими конкретный признак, а участок ДНК, в котором расположен ген данного признака.
По своему основному действию гены могут быть доминантными (обозначаются прописными буквами A, B, C, D и т. п.) и рецессивными (обозначаются соответственно строчными буквами a, b, c, d и т. п.). Ген A и его рецессивный ген а составляют пару аллельных генов данного локуса, обусловливающих определенный признак.
Доминантные гены обеспечивают проявление признаков конкретного локуса уже в первом (дочернем) поколении потомства, а рецессивный ген, полученный от другого родителя, не вызывает проявление этого признака и находится в генотипе потомка в недействующем, скрытом состоянии и может проявиться и оказать влияние только в том случае, если и отец и мать передали потомку этот рецессивный ген.
В результате слияния гамет родителей у потомка формируется генотип, т. е. набор генов обоих родителей. Если оба родителя несли доминантный ген А, то потомок будет иметь гомозиготный генотип АА с доминантным проявлением признака в фенотипе. Если оба родителя несли и передали потомку рецессивный ген а, то потомок будет гомозиготен по этому гену, его генотип будет записан аа и в фенотипе выявится рецессивный признак.
Если же от одного из родителей получен ген А, а от другого ген а, то потомок будет иметь гетерозиготный генотип Аа, а по фенотипу выявится доминантный признак. При скрещивании гетерозиготных особей между собой (Аа x Аа) у их потомства наблюдается «расщепление» по фенотипу и появляются особи как с доминантным, так и с рецессивным признаком.
Рассмотрим пример с наследованием длины шерсти у собак. Нормальная (короткошерстная) шерсть доминантна (L) над длинной шерстью (l). Если скрещивать гомозиготных короткошерстных собак (LL) с длинношерстными (ll), то их гаметы с генами L и l дадут в первом поколении (F1) гетерозиготное потомство Ll, по фенотипу оно будет короткошерстным, а по генотипу гетерозиготным.
Если далее скрещивать гетерозиготных собак (F1) между собой Ll X Ll, то во втором поколении (F2) будет иметь место расщепление как по фенотипу, так и генотипу, что видно из следующей таблицы (решетка Пеннета).
В этом примере иллюстрируются два закона Менделя:
1. Единообразие потомства первого поколения с доминантным проявлением фенотипа и гетерозиготным генотипом (Ll).
2. Расщепление во втором поколении (F2) потомства по фенотипам 3: 1, а по генотипам 1: 2: 1.
Если учесть одновременное наследование по двум при знакам (дигибридное скрещивание), то наследование будет сопровождаться увеличением изменчивости и комбинированием исходных родительских признаков у потомства. Примером этого может быть скрещивание при наличии у собак длинной шерсти (l) и черной окраски (B) (ньюфаундленд) с собакой, имеющей короткошерстность (L) и шоколадную (коричневую) окраску шерсти (b) (доберман-пинчер). Тогда наследование будет характеризоваться следующим:

Родители гомозиготные: доберман-пинчер Llbb x ньюфаундленд llBB. Все их потомки (F1) будут короткошерстные черные LlBl. Скрещивание потомков LlBb x LlBb дает в F2 следующее расщепление.
Из таблицы видно, что при дигибридном скрещивании, т. е. при двух учтенных признаках из 16 возможных вариантов фенотипов получаем следующее соотношение 9 короткошерстных черных, 3 короткошерстных коричневых, 3 длинношерстных черных, 1 длинношерстный коричневый (9: 3: 3: 1).
Приведенный пример демонстрирует 3-й закон Менделя: признаки при доминантном и рецессивном действии Генов разных локусов наследуются независимо. Такое наследование создает новые фенотипы, которых не было в предыдущих поколениях и тем самым повышается изменчивость, называемая комбинативной.
При учете наследования по трем признакам из 64 возможных получают соотношение фенотипов 27: 9: 9: 9: 3: 3: 3: 1. Такое расщепление будет получено при скрещивании гетерозиготных собак (F1) по 3 признакам: длина шерсти, цвет шерсти, сплошная (или пятнистая) окраска.
Комбинативная изменчивость используется при выведении новых пород, когда ставится цель создать у собак такие комбинации признаков, которых не было у исходных пород или помесей и которые закрепляются далее определенной системой подбора пар.
Вместе с тем явление независимо друг от друга на следования признаков и явление расщепления в потомстве гетерозиготных родителей усложняют племенную работу, так как указывают на «засоренность» породы нежелательными генами (признаками) и требуют селекционной очистки популяции от таких генов.
Кроме наследования в виде доминантности и рецессивности генов, может иметь место совместное воздействие разных аллелей данного локуса: кодоминантное действие генов. Например, синтез белка гемоглобина обусловлен генами А и В, которые дают гемоглобин трех типов АА, ВВ и АВ, и каждый из генотипов обеспечивает синтез нормальных гемоглобинов. Проявляются различия совместного действия А и В только в биохимической структуре молекулы соответствующими методами путем электрофореза образцов крови.
Кроме отмеченных закономерностей в наследовании признаков потомства, обусловленных взаимодействием аллелей одного локуса, наблюдается такая особенность, как появление нового состояния признака у потомство», которое отсутствовало у его родителей. Этот тип наследования называется «новообразованием при скрещивании». Примером такого наследования служит скрещивание кофейного (коричневого) добермана с голубым доберманом. В результате их скрещивания получаются доберманы черного цвета, так как у исходных типов доберманов различны аллели генотипа локуса. Генотип кофейного добермана включает ген D, определяющий интенсивность окраски и ген b, как рецессивный аллель гена черного цвета. Поэтому кофейный доберман имеет генотип bbDD. Генотип голубого добермана включает доминантный ген черной окраски B, но эта черная окраска не может полностью проявиться из-за отсутствия гена D (усилителя). В результате получается голубой доберман с генотипом BBdd. При скрещивании доберманов обоих типов bbDD x BBdd их потомство будет иметь гетерозиготный генотип BbDd, а по окраске все потомство будет черного цвета. От скрещивания таких гетерозиготных особей будет происходить расщепление по фенотипам в соотношении: черные 9BD+голубые 3Bd и кофейные разных оттенков 3bD+lbd, т. е. соотношение, как при обычном дигибридном скрещивании 9: 3: 3: 1.
Взаимодействие неаллельных генов (находящихся в разных участках хромосом) также приводит к новообразованию: комплементарному взаимодействию генов. При таком типе наследования расщепление по фенотипам во втором поколении будет отмечаться по их соотношению от вышеописанных. Например, в F2 может быть соотношение 9: 7 или 9: 3: 4, 12: 3: 1 при наличии разных аллелей в локусе A и локусе E. У собак соотношение 9: 7 прослежено при скрещивании гетерозиготных черных собак (генотип AsAYEe) между собой. Фенотипы их потомства были следующие: 9 черных и 7 желтых. Скрещивание черных гетерозиготных собак с генотипом As’a’Ee дает расщепление в потомстве следующего типа: 9 черных, 3 желтых, 4 рыжевато-коричневых. А при наличии других аллелей у черных гетерозиготных собак с генотипом AsAYEbrE соотношение фенотипов с новообразованием будет еще более отличающимся, а именно: 12 черных, 3 полосато-тигровых (DYDYEbrE) и 1 желтая (DYDYEE).
В наследовании, некоторых признаков проявляется действие особых генов: генов-модификаторов, влияющих на степень проявления признака. Например, они могут существенно повлиять на окрас: от сплошной окраски через серию пятнистости почти до полностью белой окраски.
Количественные признаки обусловлены влиянием многих генов. Это так называемый полигенный тип наследования, при котором действие генов приводит к тому, что количественный признак может принимать разную величину, т. е. наблюдается его варьирование от минимального до максимального значения. На фенотипическую изменчивость таких признаков оказывают существенное влияние факторы внешней среды, особенно кормление и условия содержания, но при этом сохраняется наследственная обусловленность признака. Например, высота в холке у такс варьирует у особей в пределах породы, но типичная низкорослость обусловлена наследственностью и действием многих генов.
Существенное значение в наследственности имеет плейотропное (множественное) действие гена, заключающееся в том, что один и тот же ген может влиять на разные признаки. У собак действие этого гена вызывает бесшерстность, дефекты и недоразвитие зубной системы, у борзых, например, белую окраску шерсти, глухоту. У собак породы дункер описан полулетальный * ген «крапчатости» с плейотропным действием. Он вызывает специфическую окраску шерсти в виде крапчатости, уменьшение размера глазного яблока, дефект радужной оболочки (коломбо), глаукому (повышенное глазное давление с выпячиванием глазного яблока и далее слепоту), голубую окраску радужной оболочки, глухоту, общую слабость, пониженную функцию размножения. Плейотропное действие может вызвать и развитие ценных признаков у собаки.
Кроме того, при независимом совместном наследовании генов, имеет место «сцепленное» наследование разных признаков, при котором гены «сцепленных» признаков находятся в одной и той же хромосоме и передаются через нее совместно.
На основе этого явления для некоторых видов состав лены карты хромосом, которые указывают на место расположения того или иного гена. У овец породы прекос установлено сцепление наследования крипторхизма с комолостью баранов. У собак сцепленное наследование связано с присутствием некоторых генов в половой Х-хромосоме: (гены крипторхизма (ген c), гемофилии (ген h).
Под влиянием ряда внешних факторов (рентгеновские лучи, химические вещества), а также в результате изменения обменных процессов при старении организма, в хромосомном наборе гамет и соматических клеток могут происходить перестройки хромосом, вызывающие наследственные мутационные изменения. Мутационные изменения подчас затрагивают и перестраивают химическую структуру в молекуле ДНК (генные мутации), что в свою очередь приводит к появлению нового состояния гена, т. е. его новой аллельной форме. Чаще всего исходный доминантный ген превращается в мутантный аллель.
Как правило, он бывает рецессивным, и его присутствие выявляется только в последующих поколениях. Реже происходят мутации рецессивного гена в доминантный. Подавление мутантного гена у родителя обнаружится лишь в последующих поколениях, если рецессивный мутантный аллель, например а, будет получен от обоих родителей, несущих в гаметах рецессивный аллель а, что приведет к формированию у потомков гомозиготного рецессивного состояния генотипа aa, и рецессивный ген обоих аллелей вызовет формирование нового признака (свойства), что проявится в фенотипе такого потомка.
Мутации могут происходить в виде поломок и перестроек самих хромосом, путем обмена участками между хромосомами-аналогами. В процессе мутагенеза возможно даже изменение числа хромосом в кариотипе в виде утраты или добавки отдельных хромосом или путем увеличения числа пар хромосом (полиплодия).
Мутационные процессы в кариотипе сопровождаются изменением свойств соматических клеток или гамет, в результате чего изменяется их наследственность, что сопровождается появлением новых особенностей в клетке или организме. Так, если мутация происходит в соматических клетках, это может вызвать опухоли в данной ткани. Мутации, происходящие в половых клетках родителей, приводят к изменению и появлению новых свойств у их потомства.
Мутационные изменения в большинстве случаев вызывают аномалии, уродства, болезни и гибель потомства как на первых этапах развития зародыша, так и в более поздние периоды. Если в приплоде некоторых самцов или самок регистрируются аномалии или наследственные болезни, то таких собак нельзя использовать в племенной работе. Но следует иметь в виду, что некоторые мутации можно использовать для создания новых пород.
Если у потомства нарушается нормальное число половых хромосом и в кариотипе вместо нормы XX ( ) и XY ( ) образуются наборы типа XXY, XXXY, YYX и др., это приведет к нарушению половой функции, полной половой стерильности.
Мутирование исходного доминантного аллеля может происходить многократно, в результате чего образуется серия рецессивных аллелей. Между аллельными генами формируется определенная последовательность в степени проявления признака.
По данным Робертсона (1982), серия множественного аллелизма выявлена в отношении синтеза пигмента шерсти у собак. Каждый новый аллель локуса пигментации, вызывая синтез нового пигмента, дает другую окраску, что и было использовано при выведении новых пород. Приведем серию множественных аллелей, обусловливающих пигментацию шерсти у собак.

* «Агути» проявляется в виде кольцевых зон разной окраски шерстинок по их длине.
Взаимоотношение между членами этой серии следующее (от наиболее доминантного действия в сторону уменьшения интенсивности признака):
As > Ay > A > asa > at.
Возникновение новых аллелей в мутационном процессе, вызванное естественными или искусственными воздействиями внешних факторов, может затрагивать и изменять различные признаки и свойства организма, такие как строение скелета (черепа, конечностей, туловища), размеры тела, особенности шерстного покрова по типу и окраске шерстинок, тип конституции, телосложение, инстинкты и т. д. Поэтому повышенная изменчивость, вызываемая мутированием, была использована человеком широко и удачно, особенно для выведения пород собак, отвечающих разнообразным запросам и вкусам человека.
В собаководстве наиболее подробно разработана генетика окраски шерсти и пегости со многими вариациями, генетика типа шерстинок и шерстного покрова, генетика болезней и аномалий.
Каждая порода характеризуется разной степенью изменчивости этих признаков и разными особенностями наследования.
В современной литературе (Р. Робертсон, 1982) дано описание генетических особенностей 118 пород собак разного направления, для которых выявлено 29 аллелей пигментации. Так, например, по фенотипической и генетической изменчивости мастей наибольшая вариация выявлена, у кокер-спаниеля (18 фенотипов), у пинчера (10 фенотипов), у пуделей (14 фенотипов).
Значительно меньшая изменчивость окраски шерсти наблюдается у собак сторожевого, служебного направления, несколько больше у собак охотничьих, промысловых.
У собак выявлены гены и их аллели для следующих окрасок: серая окраска (немецкая овчарка); черная (ньюфаундленд); кофейные (печеночного оттенка, коричневые) (доберман-пинчеры); голубые (доберман-пинчеры); соболиная, красная (боксеры); желтая, кремовая, светло-палевая (изабелла); чепрачные (колли); черные с подпалом (доберман-пинчеры); тигровые (доги, боксеры); альбиносы (лейцисты); арлекины.
Генетически обусловлено также распределение пигментации по телу (Н. А. Ильин). Различают собак одноцветных, двух и трехцветных (сеттеры). Пигментация может быть от сплошной, пятнистой разной степени и типа рисунка, до сильно выраженной утраты пигмента. Типы распределения пигмента сильно варьируют, а в ряде пород служат породным признаком.
Специфическую пятнистость имеют пойнтеры и сеттеры. У них кроме обычной пятнистости в виде пегости наблюдается так называемая крапчатость: мелкие штрихообразные или округлого типа черные, коричневые мелкие пятна по белому фону. У гончих, кроме обычной чепрачности черного или коричневого цвета, и у сеттеров описана «тиковая испещренность»: белые волосы равномерно перемешаны с пигментированными. Она имеет доминантное наследование (ген T). Тиковая испещренность начинает формироваться у щенка к месячному возраст.
Пигментация может распространяться по всей длине волоса (шерстинки) или в виде поперечно окрашенных разного цвета зон (зонарная). Зонарная окраска распространена у немецких овчарок, лаек и может сопровождаться чепрачным типом распределения по телу в виде тигрового (полосатость) рисунка, с подпалами.
Чалость (смесь неокрашенных (белых) волос с окрашенными) обусловлена доминантным геном ®. У догов с мышастой шерстью в генотип входит этот ген.
Вот некоторые данные, характеризующие фенотипические особенности пигментации собак разных пород.
Афганские породы: черная, черная с коричневым подпалом, голубая, серая.
Бедлингтон терьер: голубой, голубой с коричневым, печеночного цвета, песочные, кремовые.
Борзые: черные, рыжие.
Боксеры: красные, тигровые.
Бульдоги: красные, соболиные, тигровые.
Чау-чау: черные, голубые, красные, серые.
Доги: черные, коричневые, желтые, пестрые, арлекины.
Доберман-пинчеры: черные, коричневые, голубые, изабелла.
Японский хин: черные с белым, красные с белым.
Ньюфаундленды: черные; печеночного цвета, голубые.
Чи-хуа-хуа: 10 разных фенотипов.
Несмотря на большое фенотипическое разнообразие между породами и внутри их, окрас собак, как показали генетические работы, обусловлен следующими основными генами и их аллелями.
Для того чтобы в организме мог синтезироваться пигмент, влияющий на окрас шерсти, глаз, мочки носа, необходимо присутствие в генотипе гена C. Этот ген сам не вызывает окрас, а обеспечивает синтез пигмента как такового. При отсутствии доминантного гена С, при ее рецессивном аллеле с, даже при наличии аллелей, дающих черную, коричневую и другие окрасы, собаки будут иметь белую шерсть, белую мочку носа, бесцветную радужную оболочку глаза (красные глаза), т. е. будут альбиносами. Таких собак не зарегистрировано. Но в ряде пород встречается неполный альбинизм (лейцисты). Они обладают бесцветной (белой) шерстью, но сохраняют темную пигментацию мочки носа и радужной оболочки глаз. Лейцизм зарегистрирован и считается породным признаком у белых шпицев, белых бультерьеров, у некоторых пород лаек.
Следовательно, все окрашенные собаки в генотипе имеют ген C и другие гены (черный, шоколадный, желтой пигментации).
Учитывая действие указанных основных генов пигментации, приведем в качестве иллюстрации полные генотипы собак определенной окраски и породной принадлежности (по Н. А. Ильину).

С окраской шерсти коррелирует часто и пигментаци глаз. Различают карие, коричневые, желтые, голубые, голубовато-белесые, резко рубиновые (из-за отсвечивания зрачка) глаза. Иногда наблюдается разноглазость до цвету радужной оболочки у одной и той же особи, что встречается у «арлекинов».
Получили генетическое изучение и другие элементы экстерьера: тип и структуры шерстного покрова, форма и длина ушей и хвостов, особенности строения костей черепа.
Скрещивание короткошерстной собаки, несущей в генотипе доминантный ген L, с длинношерстной собакой, имеющей рецессивный ген l, дает в потомстве часто не полное, а промежуточное наследование длины шерсти В таких случаях в приплоде часто появляются особи с типом волоса, отклоняющегося в той или иной степени в сторону одного или другого родителя. Это объясняется тем, что длина волоса является количественным признаком и обусловлена влиянием многих генов, т. е. имеет полигенное наследование.
Наследование формы и размера ушной раковины так же обусловлено генетически несколькими аллелями. Установлено, что полустоячие уши (ген Ha) характерны для собак породы колли, фокстерьеров, которые могут иметь генотипы HaHa, HaH, Hah.
Висячее ухо (ген H) с генотипом HH распространено у многих пород (спаниели, гончие, таксы) и стоячее, ген с генотипом hh (немецкая овчарка), полувисячее (Hh). Часто наблюдаются промежуточные формы и размеры ушей. Наследование ушной раковины подчинено влиянию нескольких однозначных генов, т. е. имеет тип полимерного наследования. Условия выращивания и климатические факторы также могут оказывать влияние на этот признак, потому что эти факторы влияют в определенной мере на формирование конституции, появление рыхлости или, наоборот, излишней сухости и переразвитости.
Характерным наследственным и породным признаком служит длина и форма хвоста. Различают породы длиннохвостые, со средней длиной хвоста (до скакательного сустава), короткохвостые и бесхвостые (полное отсутствие хвостовых позвонков). Наследование длины хвоста обусловлено полимерией. Поэтому скрещивание длиннохвостой с короткохвостой собакой дает в их потомстве собак с варьированием длины хвоста. Однако в практике собаководства наблюдаются случаи рождения короткохвостых собак с уменьшенным числом позвонков, в результате длина хвоста у таких животных составляет около 1/3 части нормальной длины.
Считают, что на фенотипическую длину хвоста оказывают влияние гены-модификаторы и в некоторой степени внешние факторы.
Форма хвоста и его постановка наследственны и закреплены человеком путем селекции. При экстерьерной оценке собак на рингах этим особенностям придается важное значение, как элементу, характеризующему породу. Эти требования распространяются, например, на лаек (хвост кольцом на спине), фокстерьеров и эрделей (прямостоячий хвост), легавых (горизонтальное положение хвоста прутом) и др.
Большое разнообразие в строении черепа у собак часто вызывается мутацией и закрепляется селекцией, как породный признак. В результате доместикации и искусственного отбора и подбора основные морфологические особенности костей черепа, типичные для волка и дикой собаки, претерпели изменения. Удлиненные кости черепа, особенно челюстного аппарата, типичны для собак борзых пород, мощный и тяжеловесный череп характерен для ряда сторожевых собак, мопсовидность с крайним проявлением в строении черепа наблюдается у различных пород болонок, мопсов и т. п. Мутационные изменения в строении скелета конечностей (например, коротконогость), являющиеся для дикого вида собачьих пороком, были закреплены человеком в качестве породного признака у такс. Короткие ноги позволяют собаке удобнее работать в норных условиях охоты.
Прибылые пальцы также наследственно обусловленный признак. В практике собаководства их обычно удаляют у щенков в первые дни после рождения, чтобы предотвратить возможные травмы у взрослого животного.
Не будем касаться других элементов экстерьера, но заметим, что для каждой породы собак экстерьер и конституция типичны и обусловлены наследственностью, которая основывается на полигенном, плейотропном действии гена.
Наиболее важной характеристикой, на основании которой можно судить о состоянии популяции (породы), ее прогрессе и недостатках, являются наследственные болезни собак.
Наличие наследственных болезней у собак, приводящих к гибели или патологическому развитию, требует от ветеринарных врачей и кинологов тщательной регистрации дефектов. Необходимо выявление путем анализа родословных в нескольких рядах предков и у боковых родственников наличия дефектов в целях установления как явно ненормальных животных, так и скрытых носителей детальных генов в гетерозиготном состоянии, у которых в их личном фенотипе патология скрыта, а будет передаваться и выявляться у потомков.
Основой для селекционно-племенной работы кинологов, работающих в системе служебного собаководства, а также охотничьего и декоративного собаководства, должны служить генетические достижения современной науки.
Важным условием правильного проведения племенной работы в собаководстве является тщательный и правдивый учет и выявление всех отклонений от нормы морфологических, экстерьерных и физиологических качеств собак. Необходимо разъяснять владельцам собак важность правильной информации о качестве приплода, не допускать укрытия дефектов.
Следует также помнить, что для реализации и распространения в породе желательных качеств важно создавать необходимые условия кормления, воспитания и дрессировки животных, которые служат необходимым фактором, обеспечивающим реализацию генетического потенциала породы, реализацию функции так называемых «спящих» генов, улучшающих породу или влияющих на создание новых пород.
Большое значение для развития собаководства имеет квалифицированная подготовка кадров кинологов в плановом порядке через систему вузов, а также через курсы при клубах служебного собаководства.